Jaringan Spesifik pada Jalur Steroidogenesis
Jalur Adrenal
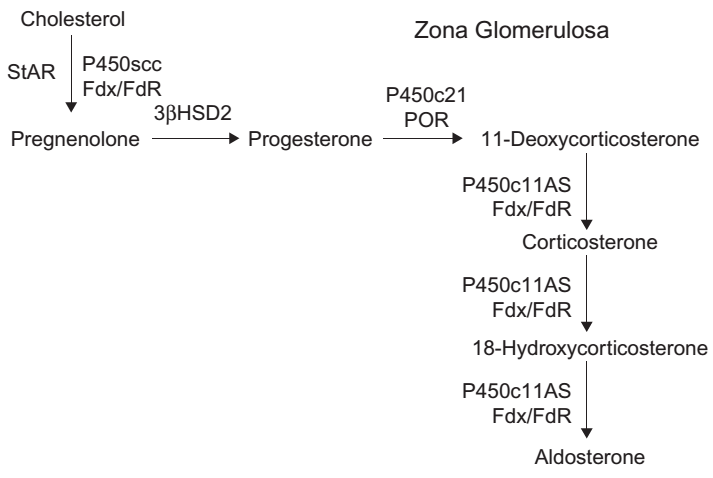
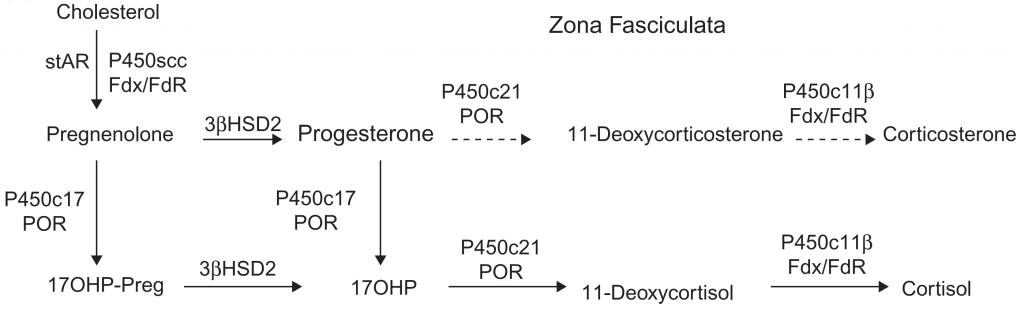
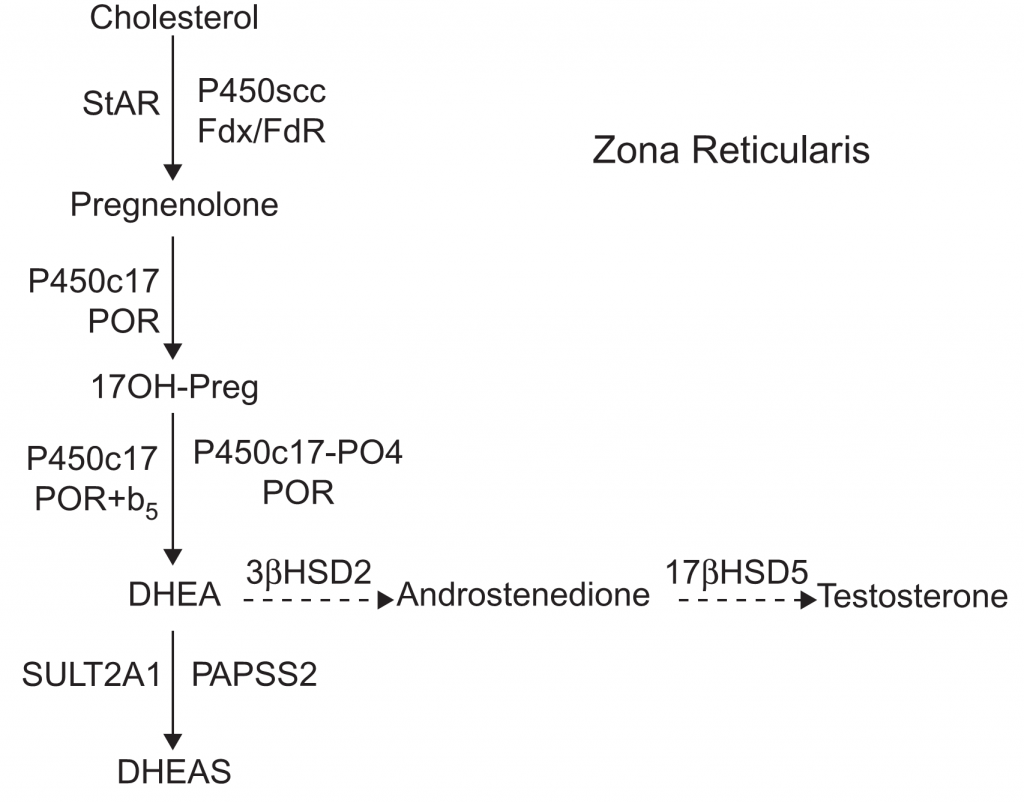
Diagram jalur steroidogenik, biasanya menggabungkan jalur dari beberapa jenis sel untuk memberikan gambaran umum dari semua proses steroidogenik. Namun diagram-diagram tersebut, tidak sepenuhnya benar, karena jalur yang terjadi pada setiap sel bisa berbeda-beda. Pada kelenjar adrenal, steroidogenesis melalui tiga jalur utama (Gambar 1, 2, & 3). Zona glomerulosa (ZG) adrenal memiliki tiga ciri yaitu mengekspresikan reseptor angiotensin II, mengekspresikan P450c11AS dan tidak mengekspresikan P450c17. Akibatnya ZG menghasilkan aldosteron di bawah regulasi oleh sisten renin/angiotensin. Sebaliknya zona fasciculata (ZF) adrenal tidak mengekspresikan reseptor angiotensin II atau P450c11AS, melainkan mengekspresikan MC2R (reseptor ACTH) dan P450c11β, yang tidak dapat mengubah 18-hydroxycorticosterone menjadi aldosterone dan memiliki kapasitas minimal untuk mengubah kortikosteron menjadi 18-hydroxycorticosterone. Baik ZG maupun ZF mengekspresikan P450c21, tetapi ZF juga mengekspresikan P450c17 sehingga memungkinkan mensintesis kortisol. ZF, mengekspresikan b5 sehingga P450c17 di ZF mengkatalisis 17α-hydroxylation tetapi dengan sedikit aktifitas 17,20- lyase. ZF menghasilkan kortisol dan kortikosteron dibawah pengaruh ACTH, tetapi sedikit DHEA. Zona retikularis (ZR) adrenal juga mengekspresikan MC2R, tetapi sangat sedikit mengekspresikan P450c21 atau P450c11β, sehingga ZR menghasilkan kortisol yang sangat sedikit. Sebaliknya ZR mengekspresikan P450c17 dan sitokrom b5 dalam jumlah besar sehingga memaksimalkan aktivitas 17,20-lyase, sehingga DHEA diproduksi yang banyak disulfasi oleh SULT2A1 menjadi DHEA. Lebih lanjut, ZR mengekspresikan relatif sedikit 3βHSD2 dan jumlah yang dibutuhkan untuk pregnenolone dan 17OH-Preg adalah 5 μM sedangkan untuk 17α-hydroxylase dan 17,20-lyase dari P450c17 membutuhkan 1 μM. Saat DHEA terakumulasi, sebagian kecil diubah menjadi androstenedion, yang kemudian sebagian kecilnya akan diubah menjadi testosteron oleh AKR1C3/17βHSD5. Oleh karena itu, pola produksi steroid yang disekresikan oleh setiap zona di adrenal ditentukan oleh enzim yang diproduksi pada zona tersebut.Diagram jalur steroidogenik, biasanya menggabungkan jalur dari beberapa jenis sel untuk memberikan gambaran umum dari semua proses steroidogenik. Namun diagram-diagram tersebut, tidak sepenuhnya benar, karena jalur yang terjadi pada setiap sel bisa berbeda-beda. Pada kelenjar adrenal, steroidogenesis melalui tiga jalur utama. Zona glomerulosa (ZG) adrenal memiliki tiga ciri yaitu mengekspresikan reseptor angiotensin II, mengekspresikan P450c11AS dan tidak mengekspresikan P450c17. Akibatnya ZG menghasilkan aldosteron di bawah regulasi oleh sisten renin/angiotensin. Sebaliknya zona fasciculata (ZF) adrenal tidak mengekspresikan reseptor angiotensin II atau P450c11AS, melainkan mengekspresikan MC2R (reseptor ACTH) dan P450c11β, yang tidak dapat mengubah 18-hydroxycorticosterone menjadi aldosterone dan memiliki kapasitas minimal untuk mengubah kortikosteron menjadi 18-hydroxycorticosterone. Baik ZG maupun ZF mengekspresikan P450c21, tetapi ZF juga mengekspresikan P450c17 sehingga memungkinkan mensintesis kortisol. ZF, mengekspresikan b5 sehingga P450c17 di ZF mengkatalisis 17α-hydroxylation tetapi dengan sedikit aktifitas 17,20- lyase. ZF menghasilkan kortisol dan kortikosteron dibawah pengaruh ACTH, tetapi sedikit DHEA. Zona retikularis (ZR) adrenal juga mengekspresikan MC2R, tetapi sangat sedikit mengekspresikan P450c21 atau P450c11β, sehingga ZR menghasilkan kortisol yang sangat sedikit. Sebaliknya ZR mengekspresikan P450c17 dan sitokrom b5 dalam jumlah besar sehingga memaksimalkan aktivitas 17,20-lyase, sehingga DHEA diproduksi yang banyak disulfasi oleh SULT2A1 menjadi DHEA. Lebih lanjut, ZR mengekspresikan relatif sedikit 3βHSD2 dan jumlah yang dibutuhkan untuk pregnenolone dan 17OH-Preg adalah 5 μM sedangkan untuk 17α-hydroxylase dan 17,20-lyase dari P450c17 membutuhkan 1 μM. Saat DHEA terakumulasi, sebagian kecil diubah menjadi androstenedion, yang kemudian sebagian kecilnya akan diubah menjadi testosteron oleh AKR1C3/17βHSD5. Oleh karena itu, pola produksi steroid yang disekresikan oleh setiap zona di adrenal ditentukan oleh enzim yang diproduksi pada zona tersebut.Diagram jalur steroidogenik, biasanya menggabungkan jalur dari beberapa jenis sel untuk memberikan gambaran umum dari semua proses steroidogenik. Namun diagram-diagram tersebut, tidak sepenuhnya benar, karena jalur yang terjadi pada setiap sel bisa berbeda-beda. Pada kelenjar adrenal, steroidogenesis melalui tiga jalur utama. Zona glomerulosa (ZG) adrenal memiliki tiga ciri yaitu mengekspresikan reseptor angiotensin II, mengekspresikan P450c11AS dan tidak mengekspresikan P450c17. Akibatnya ZG menghasilkan aldosteron di bawah regulasi oleh sisten renin/angiotensin. Sebaliknya zona fasciculata (ZF) adrenal tidak mengekspresikan reseptor angiotensin II atau P450c11AS, melainkan mengekspresikan MC2R (reseptor ACTH) dan P450c11β, yang tidak dapat mengubah 18-hydroxycorticosterone menjadi aldosterone dan memiliki kapasitas minimal untuk mengubah kortikosteron menjadi 18-hydroxycorticosterone. Baik ZG maupun ZF mengekspresikan P450c21, tetapi ZF juga mengekspresikan P450c17 sehingga memungkinkan mensintesis kortisol. ZF, mengekspresikan b5 sehingga P450c17 di ZF mengkatalisis 17α-hydroxylation tetapi dengan sedikit aktifitas 17,20- lyase. ZF menghasilkan kortisol dan kortikosteron dibawah pengaruh ACTH, tetapi sedikit DHEA. Zona retikularis (ZR) adrenal juga mengekspresikan MC2R, tetapi sangat sedikit mengekspresikan P450c21 atau P450c11β, sehingga ZR menghasilkan kortisol yang sangat sedikit. Sebaliknya ZR mengekspresikan P450c17 dan sitokrom b5 dalam jumlah besar sehingga memaksimalkan aktivitas 17,20-lyase, sehingga DHEA diproduksi yang banyak disulfasi oleh SULT2A1 menjadi DHEA. Lebih lanjut, ZR mengekspresikan relatif sedikit 3βHSD2 dan jumlah yang dibutuhkan untuk pregnenolone dan 17OH-Preg adalah 5 μM sedangkan untuk 17α-hydroxylase dan 17,20-lyase dari P450c17 membutuhkan 1 μM. Saat DHEA terakumulasi, sebagian kecil diubah menjadi androstenedion, yang kemudian sebagian kecilnya akan diubah menjadi testosteron oleh AKR1C3/17βHSD5. Oleh karena itu, pola produksi steroid yang disekresikan oleh setiap zona di adrenal ditentukan oleh enzim yang diproduksi pada zona tersebut.
Jalur Gonad
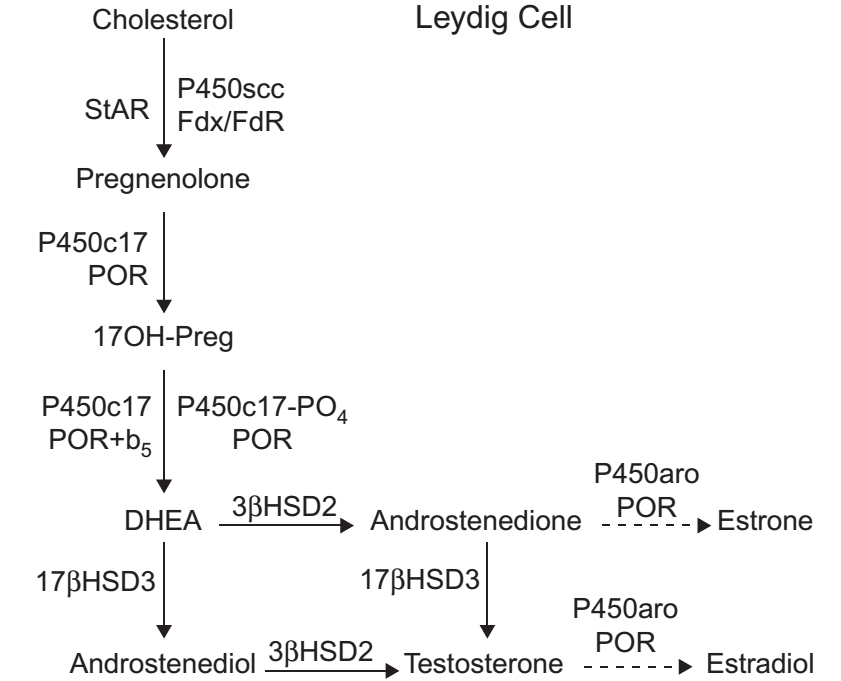
Sintesis testosteron di testis mengikuti jalur yang mirip dengan jalur produksi C1-steroid di ZR adrenal, dengan pengecualian penting bahwa stimulus untuk steroidogenesis ditransduksi oleh reseptor LH daripada MC2R dan bahwa sel Leydig mengekspresikan 3βHSD2 dan 17βHSD3, tetapi tidak mengekspresikan SULT2A1. Jadi DHEA yang diproduksi di testis tidak tersulfasi tetapi lebih mudah menjadi androstenedion dan testosteron (Gambar 4). Seperti pada jalur adrenal, jalur utama steroid C19 adalah melalui Δ5 steroids ke DHEA, jalur Δ4 dari 17OHP ke androstenedion hanya memberikan kontribusi yang kecil.
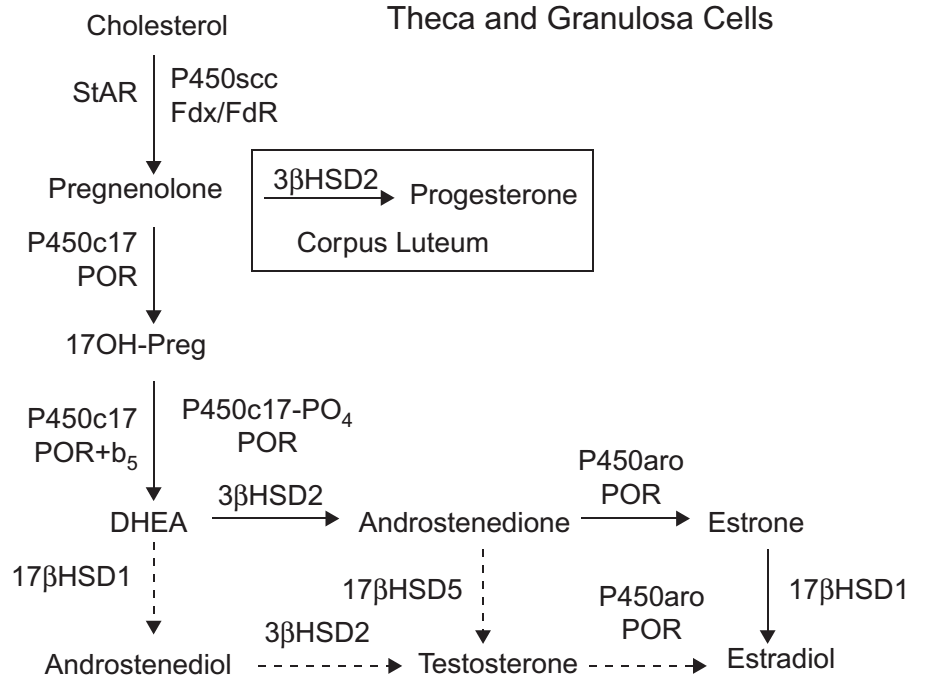
Steroidogenesis di ovarium terbagi antara granulosa dan sel teka, yang mengelilingi oosit dan membentuk folikel. Pola steroidogenesis juga bervariasi selama siklus menstruasi. Estradiol adalah produk utama dalam fase folikuler sedangkan progesteron diproduksi pada fase luteal (Gambar 5). Poin kunci dalam steroidogenesis pada ovarium adalah bahwa sel granulosa tidak mengekspresikan P450c17. Jadi secara umum, steroidogenesis dimulai dalam sel granulosa dibawah pengaruh LH, melalui cAMP yang merangsang ekspresi P450cc. Pregenolone dan progesteron dari sel granulosa kemudian berdifusi ke dalam sel teka yang berdekatan, yang kemudian mengubah androstenedion oleh P450c17 dan 3βHSD2. Sejumlah kecil androstenedion ini disekresikan atau diubah menjadi testosteron, tetapi sebagian besar androstenedion kembali ke sel granulosa dimana akan diubah menjadi estron dan kemudian menjadi estradiol oleh P450aro dan 17βHSD1 dibawah pengaruh hormon FSH. Jadi seperti pada tiga zona adrenal, pola steroidogenesis pada gonad di ovarium ditentukan oleh ekspresi spesifik dari enzim steroidogenik spesifik.
Regulasi Kronis Steroidogensis
Jika StAR merupakan penentu regulasi akut steroidogenesis, P450scc adalah langkah pembatas laju enzimatik dalam steroidogenesis. Dengan demikian, regulasi steroidogenesis secara kuantitatif (berapa banyak steroid yang diproduksi) ditentukan oleh ekspresi gen P450scc dan secara kualitatif (steroid mana yang diproduksi) ditentukan oleh enzim hilir terutama P450c17. Seseorang dengan mutasi yang menginaktivasi reseptor ACTH (MC2R) atau LH/hCG, menyebabkan steroid dapat diabaikan dari adrenal atau gonad. Sebaliknya, mengaktifkan mutasi reseptor LH menyebabkan hipersekresi steroid dan elemen responsif cAMP telah diidentifikasi dalam gen untuk sebagian besar enzim P450 steroidogenik manusia. Namun regulasi pembentukan cAMP saja tidak menjelaskan keragaman produksi steroid yang diamati di berbagai zona korteks adrenal dan gonad dari kedua jenis kelamin. Faktor transkripsi lainnya seperti AP2, SP1, SP3, NF1C, NR4A1, NR4A2, GATA4, and GATA6, juga berpartisipasi dalam mengatur transkripsi dan cAMP daris etiap gen. SF1 mengkoordinasikan ekspresi enzim steroidogenik di adrenal dan gonad. Sebaliknya, steroidogenesis di otak dan plasenta tidak bergantung pada SF1. Dengan demikian, perkembangan organ steroidogenik terkait erat dengan kapasitas untuk memproduksi steroid dan berbagai faktor yang bekerja pada gen untuk enzim steroidogenik untuk menghasilkan fiitur dan keragaman yang sama di antara jaringan steroidogenik.
Regulasi Akut Steroidogensis
Tidak seperti sel yang menghasilkan hormon polipeptida dan menyimpan hormon tersebut dalam jumlah besar dalam vesikula sekretorik yang siap untuk dilepaskan dengan cepat, sel steroidogenik menyimpan sangat sedikit steroid. Jadi, respon steroidogenik yang cepat memerlukan sintesis steroid baru yang cepat pula. ACTH mendorong pertumbuhan sel steroidogenik dan mempertahankan kerja steroidogenik pada tiga tingkatan yang berbeda. Pertama, bekerja dalam waktu yang cukup lama (berbulan-bulan), ACTH meningkatkan pertumbuhan adrenal, terutama sintesis IGF-2 yang dirangsang ACTH, faktor pertumbuhan fibroblast dasar, dan EGF. Faktor pertumbuhan ini kemudian merangsang hipertrofi dan hiperplasia sel adrenal dan dengan demikian, mengatur jumlah jaringan steroidogenik. Kedua, bekerja selama berhari-hari, ACTH melalui jalur cAMP dan angiotensin II melalui jalur kalsium menginisiasi transkripsi gen yang mengkode berbagai enzim steroidogenik dan protein co-faktor, sehingga menentukan jumlah produk steroidogenik dalam sel. Ketiga, ACTH dengan cepat merangsang transkripsi gen StAR dan fosforilasi pasca-translasi dalam StAR yang ada untuk meningkatkan aliran kolesterol dari membran luar ke membran dalam mitokondria untuk menjadi substrat P450scc.
Beberapa steroidogenesis adrenal terjadi terlepas dari StAR. Ketika sel non-steroidogenik ditransfeksi dengan StAR dan sistem P450scc, mereka mengubah kolesterol menjadi pregnenolone pada ~14% dari laju yang diinduksi StAR. Lebih lanjut, plasenta menggunakan mitokondria P450scc untuk menginisiasi steroidogenesis tetapi tidak mengekspresikan StAR. Mekanisme spesifik dari steroidogenesis StAR tidak jelas, dapat terjadi tanpa pemicu, atau beberapa protein dapat menggunakan aktivitas StAR untuk meningkatkan aliran kolesterol tanpa melibatkan StAR secara langsung. Mekanisme dari aksi StAR juga kurang jelas, tetapi diketahui bahwa StAR bekerja pada membran luar mitokondria, tanpa perlu memasuki mitokondria untuk menjadi aktif, dan mengalami konformasi pada membran luar sesuai dengan kebutuhan aktivitasnya. Protein ini berinteraksi untuk memindahkan kolesterol dari membran mitokondria luar ke P450scc, dan artinya kolesterol dimasukkan ke dalam membran luar masih belum jelas.