Evolusi Sel dan Sejarah Bumi: Statis dan Revolusi
Organisme hidup dapat dipelajari dengan lebih baik daripada mempelajari fosil. Bahkan tanpa fosil, studi filogenetik saat ini mampu memberikan rekonstruksi hubungan evolusioner yang sangat rinci dan akurat dari seluruh kehidupan dan menyimpulkannya. Namun, merekonstruksi sejarah kehidupan membutuhkan integrasi wawasan paleontologi dan neontologi (ilmu perbandingan organisme yang masih ada). Tanpa bukti paleontologis, pohon filogenetik tidak memiliki skala waktu sehingga kesimpulan akan kekurangan pengetahuan langsung tentang kelompok yang sama sekali punah. Bukti biologis molekuler dan sel jauh lebih sesuai dengan catatan fosil, tergantung dari penafsiran neontologis dan paleontologis. Steranes secara umum digunakan sebagai biomarker untuk identifikasi eukariota, karena beberapa bakteri secara filogenetik dapat membuat sterol, termasuk beberapa actinobacteria yang dianggap sebagai nenek moyang dari eukariota dan archaebacteria. Pemahaman terkait evolusi eukariota awal telah meningkat pesat sejak saat itu, terutama pembuktian sebagai asal usul kehidupan.
Steranes have an androstane skeleton with a side chain at carbon C-17. The sterane structure constitutes the core of all sterols.
Informasi
Cavalier-Smith menempatkan bakteri dalam bagian topologi pohon kehidupan sebagai dasar yang kuat dan menjelaskan bagaimana transisi di dalamnya dapat dipolarisasikan dengan yakin oleh argumen evolusioner yang kuat. Penulis menyimpulkan bahwa akar pohon kehidupan ada di dalam negibacteria: Eubacteria Gram negatif dengan dua pembungkus membran. Pada Interpretasi itu, membran tunggal pada posibacteria, archaebacteria, dan eubacteria adalah produk evolusi dari selubung negibacteria yang kehilangan membran luarnya. Bahwa sel pertama yang muncul adalah negibacteria dengan selubung ganda dan bahwa negibacteria kehilangan membran luarnya dalam sejarah bukanlah ide baru.
Dasar Fundamental Keanekaragaman Sel
Perbedaan Kontras antara Neomura dan Eubacteria
Dikotomi fenotipik dan genetik tunggal terbesar di dunia saat ini adalah antara prokariota dan eukariota, tetapi dikotomi yang sebelumnya muncul dan lebih mendasar adalah eubacteria (negibacteria & posibacteria) dan neomura (archaebacteria & eukariota). Transisi diantara eubacteria dan neomura melibatkan perubahan besar pada permukaan sel dan jenis protein terkonjugasi yang disintesis secara co-translasi, yaitu selama sintesis protein itu sendiri. Pada semua sel, protein membran atau semua protein yang disekresikan oleh ribosom yang melekat pada membran oleh partikel ribonukleoprotein yang termasuk jenis SRP (Signal Recognition Particle). SRP mungkin telah berevolusi bahkan sebelum terbentuk sel pertama dan dilestarikan disepanjang hidup sel, tetapi pada neomura lebih kompleks daripada eubacteria. Ini merupakan salah satu alasan yang mempertimbangkan bahwa neomura lebih maju dan telah berevolusi dari eubacteria, dan bukan sebaliknya. Neomura dan eubacteria juga sangat berbeda dalam enzim yang bekerja pada DNAnya (baik dalam replikasi, perbaikan, maupun transkripsi gen). Perbedaan dalam enzim replikasi terkadang digunakan untuk menunjukkan bahwa replikasi DNA terlibat secara independen dalam eubacteria dan neomura.
Terlepas dari perbedaan mencolok antara eubacteria dan neomura, sebagian besar sifat sel tidak begitu berbeda, dimana pada dasarnya kedua kelompok sel ini memiliki lebih dari seribu karakter yang serupa. Nenek moyang kedua sel ini adalah sel yang kompleks dan sangat berkembang, yaitu eubacterium. Dua inovasi seluler utama yang muncul pada leluhur archaebacterium yaitu flagela yang stabil terhadap asam dan lipid eter isoprenoid yang stabil terhadap asam dan panas, menunjukkan bahwa sel ini beradapatasi terhadap hipertermofili. Flagela unik sel ini berevolusi dari protein sekresi type IV yang sebelumnya digunakan untuk membentuk vili eubacterial. Saat eubacteria membuat isoprenoid, hal baru yang diperoleh archaebacteria adalah menambahkan gliserol fosfat dengan stereokimia baru untuk membuat lipid gliserofosfat isoprenoid-eter, yaitu enzim yang ditemukan hanya pada posibacteria.
Tingkat evolusi protein baru asa eukariota jauh lebih besar dibandingkan dengan archaebacterial. Perbedaan keduanya dari bakteri lain sangat dibesar-besarkan oleh perbedaan kuantitatif dalam rRNA, protein ribosom, dan enzim penanangan DNA. Persepsi archaebacteria berdasarkan perbedaan kuantitatif rRNA, sangat menyesatkan. Dua subfilum archaebacteria (euryachaeota dan Crenachaeota) mungkin merupakan nenek moyang respirator anaerob yang sangat bergantung pada senyawa belerang dengan fisiologi yang jauh lebih kuno dibanding eubacteria. Dengan menggabungkan karakter hyperthermophily, archaebacteria memperluas biosfer dengan cara yang relatif kecil ke dalam lingkungan yang panasatau asam yang sebelumnya tidak dapat diakses oleh mahluk hidup.
Perbedaan Kontras antara Negibacteria dan Posibacteria
Perbedaan paling mendasar dalam eubacteria adalah pada Posibacteria selalu dibatasi oleh satu membran (misalnya Bacillus dan Streptomuces), dan Negibacteria selalu memiliki dua membran yang berbeda (misalnya Escherichia, Salmonella, Cyanobacteria, Spirochaetes, dll). Membran dalam Negibacteria homolog dengan membran sitoplasma tunggal dari posibacteria dan archaebacteria, sedangkan membran luarnya unik dan homolog dengan membran luar mitokondria dan kloroplas. Semua lipid pada membran luar dibuat di membran dalam dan diekspor ke membran luar. Protein membran luar dibuat oleh ribosom pada permukaan sitosol membran dalam dan diangkut melintasi ruang periplasma (ruang antara membran dalam dan membran luar). Mekanisme ekspor lipid dan protein ini homolog untuk semua negibacteria, tetapi dapat diamati dengan baik hanya pada proteobacteria. Pada Negibacteria, dinding murein peptidoglikan, jauh lebih tipis daripada posibacteria dan terletak antara membran dalam dan membran luar, dan dilekatkan oleh lipoprotein.
Perbedaan Kontras antara Eobacteria dan Glycobacteria
Ciri khas Negibacteria (misalnya Proteobacteria dan cyanobacteria) memperkuat membran luar nya dengan memasukkan molekul Lipopolisakarida (LPS) yang sangat kompleks ke dalam lapisan ganda terluar dan bukan berupa fosfolipid sederhana. Enam filum negibacteria memiliki LPS tetapi hilang pada sphingobacteria, spirochaetae, dan glycobacteria. Glycobacteria juga menggunakan hopanoid untuk memperkuat membran dalamnya. Dua filum negibacteria, Chlorobacteria dan hadobacteria, yang secara seragam kekurangan LPS dan hopanoid dikelompokkan ke dalam Eobacteria. Pembungkus sel eobacteria lebih sederhana dengan saluran tubular yang unik untuk memediasi sekresi protein tipe I di seluruh membran dalam dan terlibat dalam impor molekul di seluruh membran luar.
Pohon Kehidupan dan Akarnya
Urutan pohon kehidupan menggunakan molekul tunggal seperti DNA, RNA, dll, sangat penting untuk mengelompokkan organisme yang relatif terkait erat, tetapi terdapat tiga hambata utama yaitu tidak cukupnya data untuk membentuk pohon kehidupan, beberapa gen terkadang dapat ditransfer secara lateral diantara organisme yang tidak terkait, dan bias sistematis dalam tingkat dan mode evolusi. Analisis kladistik dari karakter morfologi atau makromolekul yang cukup kompleks untuk mengesampingkan konvergensi, seringkali merupakan cara yang paling dapat diandalkan untuk membangun hubungan, meskipun dapat diganggu oleh kehilangan komponen akibat peristiwa evolusi. Analisis kladistik karakter genetik (misalnya transfer gen, penggantian gen, atau fusi gen) juga penting. Praktik filogenetik terbaik mencari kesesuaian antara semua jenis bukti. Seseorang harus mencoba memahami alasa untuk setiap konflik dan biasa dan memberikan bobot lebih pada bukti terkuat dan metode yang paling dapat diandalkan.
Negibacteria Mendahului Posibacteria
Transisi antara negibacteria dan posibacteria terjadi pada sel dengan flagela eubacteria. Menentukan apakah awalnya flagela berevolusi di posibacteria atau di negibacteria akan mempolarisasi proses transisi. Flagela eubacteria memiliki tiga bagian yaitu motor statis yang tertanam di membran dalam (biasanya secara kovalen melekat pada dinding murein), badan basal silinder (diputar oleh arus proton melalui motor terkait), dan poros heliks (diluar membran dalam dan secara pasit mentransmisikan rotasi kelingkungan). Karena prekursor evolusioner tubuh basal dan motorik hanya ada di negibacteria, dan tidak terdapat pada posibacteria, flagela berevolusi pertama kali di negibacteria dan ditransmisikan secara vertikal ke posibacterium pertama. Hal ini jelas mempolarisasi evolusi dari negibacteria ke posibacteria, dan bukan sebaliknya, sehingga ancestor posibacterium kehilangan membran luarnya. Hilangnya Membran luarmemiliki dua konsekuensi co-evolusi yaitu hilangnya cincin flagella pada negibacteria kecuali spirochaetes menanamkan tubuh basal di membran luar, dan asal enzim sortase posibacterial untuk menempelkan protein periplasmic ke dinding murein tebal.
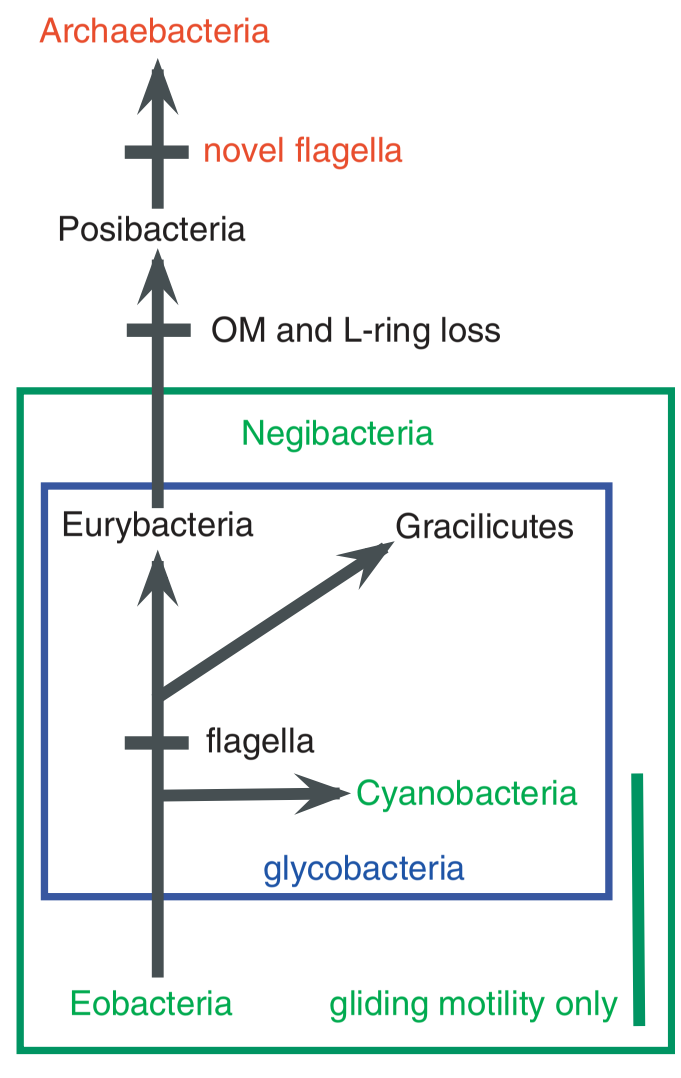
Archaebacteria Merupakan Saudara Eukariota (Bukan Ancestor)
Meskipun terkadang diasumsikan bahwa archaebacteria merupakan nenek moyang dari eukariota, bukti yang luas dan beragam sangat mendukung holophyly archaebacteria (saudara eukariota), dan idak ada konsekuensi apapun yang mendukung paraphyly (menjadi nenek moyang) dari archaebacteria. Berbagai data, termasuk sekuen RNA Ribosom menunjukkan bahwa eukariota sebagai saudara archaebacteria. Holophyly archaebacteria sangat terkait oleh kandungan lipid eter isoprenoid pada keduanya. Jika nenek moyang bersama antara archaebacteria dan eukariota memiliki lipid seperti itu, tetapi tidak memiliki lipid asil ester (ada pada seluruh eubacteria), maka nenek moyang eukariota harus memperoleh kembali lipid asil ester dan benar-benar tidak memiliki eter isoprenoid. Karena nenek moyang eukariota sudah memiliki mitokondria, sebuah protoeukariota dengan haya lipid eter isoprenoid secara teoritis dapat menggantikan seluruhnya dengan lipid asil ester, walaupun secara evolusi sangat tidak mungkin. Kelemahan lebih lanjut dari hipotesis itu adalah bahwa proteobacteria kekurangan phosphatidylinositol (salah satu jenis fosfolipid eukariota yang paling penting, diperlukan untuk pensinyalan sel eukariota spesifik). Satu-satunya eubacteria yang memiliki phosphatidylinositol adalah Actinobacteria, yang merupakan nenek moyang atau saudara dari neomura. Dengan demikian, lipid membran eukariota mungkin berasal secara vertikal dari nenek moyang Actiobaterial, lipid archaebacterial yang berasal dari nenek moyang mereka terpisah dari eukariota. Sehingga dapat disimpulkan bahwa Archaebacteria, eukariota, dan neomura memiliki hubungan holophyletic.
Neomura Berevolusi dari Actinobacteria
Eubacteria merupakan monofiletik dengan nenek moyang sama yang memiliki dinding murein peptidoglikan dan lipoprotein. Neomura juga monofiletik dengan nenek moyang sama yang memiliki N-Linked Glikoprotein, tidak memiliki murein atau lipoprotein, dan hampir pasti memiliki asil ester, tetapi tidak memiliki lipid membran eter isoprenoid. Nenek moyang ini hanya memiliki satu membran, seperti Posibacteria. Kemungkinan asal membran independen di eubacteria dan neomura yang memiliki karakter yang sama. Kunci penting hubungan neomura dan actinobacteria adalah adanya proteasome (ruang kecil untukpencernaan protein). Eubacteria lainnya tidak memiliki homolog atau yang lebih sederhana dari ini. Jadi, neomura dan actinobacteria merupakan cabang; akar pohon kehidupan diantara eubacteria lainnya.
Asal Usul Eukariota
Asal usul eukariota melibatkan perubahan paling radikal dalam struktur sel dan mekanisme pembelahan dalam sejarah kehidupan. Ini adalah contoh utama dari evolusi kuantum dengan laju evolusi yang dipercepat secara episodik secara dramatis dengan melibatkan banyak gen dan pembuatan gen baru secara besar-besaran melalui duplikasi, divergensi, dan pembentukan domain protein. Ratusan duplikasi gen dan divergensi fungsional terjadi (misalnya pada miosin, tubulin, dan protein vesikel berlapis), sehingga tidak mungkin terjadi evolusi dari eukariota ke prokariota, membalikan diferensiasi sitologi kompleks. Intron spliceosomal yang masuk dalam ribuan gen dapam cenancestor aukariota, tidak benar-benar terbalik, memskipun ada pengurangan menjadi tiga di trypanosome. Beberapa ahli biologi terkemuka yakin bahwa intron spliceosomal prokariotik adalah keturunan sekunder.
Archaebacteria Adalah Prokariota Muda
Archaebacteria dinamai ketika semua archaebacteria yang diketahui adalah metanogen (menghasilkan metana). Alasan utama mengatakan archaebacteria lebih kuno adalah bahwa rRNAnya tampak berbeda dari eukariota dan eubacteria. Stackebrandt & Woese secara eksplisit menyatakan tidak ada bukti bahwa rRNA berevolusi pada tingkat yang sama dalam tinga kelompok sehingga umumnya diabaikan.
awesomeeeee