Perkembangan Pengetahun Tentang Sel
Sejarah penemuan Sel
Robert Hooke adalah ilmuwan lulusan Oxford yang menghabiskan karirnya di Royal Society and Gresham College. Penelitian dan eksperimennya berkisar dari astronomi, biologi, hingga fisika. Pada bidang fisika, Robert Hooke diakui sebagai pencetus Hukum Hooke untuk elastisikas penggunaan mikroskop. Publikasi Hooke yang paling penting dalam Micrographia pada tahun 1665 adalah mendokumentasikan eksperimen yang dibuatnya menggunakan mikroskop. Dalam studinya, ia menciptakan istilah Sel saat membahas struktur gabus. Dia juga menggambar`kan lalat, bulu dan kepingan salju, dan dengan tepat mengidentifikasi fosil sebagai sisa-sisa mahluk yang pernah hidup.
Dalam Micrographia, Hooke menulis:
“….. Saya dapat dengan sangat jelas melihat strukturnya berlubang dan keropos, seperti sarang lebah, tetapi pori-porinya tidak teratur …. pori-pori ini, atau sel …. memang pori-pori mikroskopis pertama yang pernah saya lihat dan mungkin yang pernah terlihat, karena saya belum pernah bertemu dengan penulis atau orang manapun yang pernah menyebutkannya sebelum ini ….”
Hooke telah menemukan sel tumbuhan. Lebih tepatnya, apa yang dilihat Hooke adalah dinding sel dalam jaringan gabus. Faktanya, Hooke-lah yang menciptakan istilah “Sel”. Hooke juga melaporkan melihat struktur serupa di kayu dan tanaman lain. Pada tahun 1678, setelah Leeuwenhoek menulis kepada Royal Society dengan laporan penemuan “animalcule“, Hooke diminta oleh lembaga tersebut untuk mengkonfirmasi penemuan Leeuwenhoek. Hooke berhasil melakukannya, sehingga membuka jalan bagi penerimaan yang lebih luas terkait penemuan Leeuwenhoek. Hooke mencatat bahwa mikroskop sederhana Leeuwenhoek memberikan gambar yang lebih jelas daripada mikroskopnya.
Pada tahun 1676, Anthony van Leeuwenhoek mengamati air dari dekat dan terkejut melihat organisme kecil – bakteri pertama yang diamati oleh manusia. Suratnya yang mengumumkan penemuan ini menyebabkan keraguan yang meluas di Royal Society tetapi Robert Hooke kemudian mengulangi percobaan tersebut dan dapat mengkonfirmasi penemuannya. Selain sebagai bapak mikrobiologi, van Leeuwenhoek meletakkan dasar-dasar anatomi tumbuhan dan menjadi ahli reproduksi hewan. Dia menemukan sel darah dan nematoda mikroskopis, dan mempelajari struktur kayu dan kristal. Dia juga membuat lebih dari 500 mikroskop untuk melihat objek tertentu. Dia juga menemukan sperma, yang dia anggap sebagai salah satu penemuan terpenting dalam karirnya, dan menggambarkan spermatozoa dari moluska, ikan, amfibi, burung, dan mamalia, sampai pada kesimpulan baru bahwa pembuahan terjadi ketika spermatozoa menembus sel telur.
Makalah Leeuwenhoek pada tahun 1677 tentang protozoa memberikan deskripsi terperinci tentang protista dan bakteri yang hidup diberbagai lingkungan. Para ilmuwan pada masa itu kemudian tidak dapat menandingi resolusi dan kejelasan mikroskop Leeuwenhoek, sehingga penemuannya diragukan atau bahkan diabaikan, sehingga pengaruhnya terhadap perkembangan ilmu biologi tidak dapat diabaikan. van Leeuwenhoek menyebut para binatang kecil itu dengan istilah “animalcules” yang berarti hewan yang berukuran kecil.
Leeuwenhoek menuelidiki kembali spermatozoa dari berbagai organisme sehat (manusia, ikan, burung, cacing, anjing) dan menemukan bahwa dirinya cukup yakin untuk menulis bahwa animalcules atau spermatozoa terdiri dari begitu banyak bagian sebagai penyusun tubuh. Leewenhoek lebih lanjut berspekulasi bahwa animalcule menempelkan dirinya ke pembuluh darah di dalam rahim, dimana ia menerima makanan untuk tumbuh. Saat itu Leeuwenhoek percaya bahwa animalcules dalam bentuk spermatozoa akan menjadi sosok manusia yang dilengkapi dengan jantung dan usus lainnya, dan memang memiliki semua kesempurnaan manusia.
Teori preformasionis menyatakan bahwa sel germinal dari setiap organisme mengandung miniatur dewasa yang belum terbentuk dan akan tumbuh selama masa perkembangan embrio. Pengamatan pertama tentang sperma dilakukan oleh Anthony van Leeuwenhoek pada tahun 1677 yang melihat ribuan mahluk mirip cacing dan diberi nama animalcule. Temuan van Leeuwenhoek mendukung pandangan preformasionist. Pandangan preformasionist van Leeuwenhoek mendekati sudut pandang Aristoteles sebagai pencetus teori, karena sperma mengandung semua input yang diperlukan untuk perkembangan, sedangkan betina hanya menyediakan materi untuk pertumbuhan embrio.
Sebuah catatan kaki dalam buku karya Aristoteles yang berjudul “De Generatione Animalium” menjelaskan bahwa epigenesis berkaitan dengan pertanyaan sentral dalam perkembangan embriologis. Perdebatan antara epigenesis dan preformasionsit memunculkan kontroversi berabad-abad, yang akhirnya berpuncak pada uji coba untuk mensintesis keduanya. Untuk memahami sifat dari sintesis ini, penting untuk menyelidiki esensi dari istilah pusat epigenesis dan preformasionist. Epigenesis dalam pandangan Aristoteles adalah sebuah hierarki bentuk, dimana produk dari satu generasi bertindak sebagai materi untuk pembentukan tingkat organisasi berikutnya.
Swammerdam adalah ahli serangga yang banyak meneliti perihal metaborfosis. Salah satu penelitian yang menggugah adalah penelitian terkait dengan metamorfosis katak, dimana Swammerdam menghubungkan serangga dan katak melalui demonstrasi bahwa semuanya mengalami epigenesis. Hal ini menunjukkan kekecewaan Swammerdam akan paham preformasionist yang mengatakan bahwa hewan yang muncul diawal perkembangan menunjukkan bentuk akhir dari hewan tersebut. Swammerdam sendiri mengemukakan teori metamorfik dan epigenetik.
Dutrochet belajar kedokteran di Paris sekitar tahun 1806 dan kemudian menjabat sebagai perwira medis militer di Spanyol sebelum kemudian meninggalkan praktik kedokterannya untuk mengabdikan karirnya pada penelitian ilmiah. Ketika Dutrochet memperbatikan kesamaan proses fisik dan kimia pada hewan dan tumbuhan, ia mengarahkan studinya ke fisiologi tumbuhan dan hewan. Dutrochet adalah orang pertama yang menyelidiki secara menyeluruh mekanisme respirasi, sensitivitas cahaya, dan geotropisme pada tumbuhan, serta eksperimen klasiknya pada osmosis memperoleh pengakuan perannya dalam transportasi tanaman internal dan difusi melalui membran semipermeabel. Dutrochet juga mengungkapkan bahwa jamur adalah tubuh reproduksi dan merupakan orang yang menyatakan pentingnya sel individu dalam fungsi suatu organisme.
Robert Brown merupakan ahli botani dari Skotlandia yang terkenal karena deskripsinya tentang inti sel dan gerakan kontiniu berupa partikel kecil dalam larutan yang kemudian disebut dengan gerak Brown. Pada tahun 1827, Brown mengamati dengan menggunakan mikroskop, bahwa partikel kecil yang dikeluarkan dari butiran serbuk sari yang tersuspensi ke dalam air melakukan semacam gerakan terus menerus yang disebut dengan Gerak Brown. Dia kemudian mengamati gerakan yang sama dalam materi anorganik dan menyimpulkan bahwa gerakan itu bukan karena kekuatan tertentu.
Hugo von Mohl merupakan ahli botani berkebangsaan Jerman yang terkenal karena penelitiannya tentang anatomi dan fisiologi sel tumbuhan. Von Mohl mengembangkan gagasan bahwa inti sel berada dalam butiran bahan koloid yang membentuk substansi utama sel. Pada tahun 1846, ia menamai zat ini dengan nama “protoplasma”, sebuah kata yang ditemukan oleh ahli fisiologi Ceko, Jan Evangelista Purkinje dengan mengacu pada materi embrionik yang ditemukan dalam telur. Von Mohl juga pertama kali mengusulkan bahwa sel-sel baru dibentuk oleh pembelahan sel, sebuah proses yang ia amati pada sejenis alga. Tahun 1851, ia mengusulkan pandangan yang sekarang telah dikonfirmasi bahwa dinding sekunder sel tumbuhan memiliki struktur yang berserat.
Teori sel klasik diusulkan oleh Theodor Schwann pada tahun 1839. Ada tiga bagian dari teori ini. Pertama menyatakan bahwa semua organisme terbuat dari sel. Kedua menyatakan bahwa sel adalah unit dasar kehidupan. Bagian ini didasarkan pada kesimpulan yang dibuat oleh schwann dan Matthias Schleiden pada tahun 1838, setelah membandingkan pengamatan mereka terhadap sel tumbuhan dan hewan. Ketiga, menegaskan bahwa sel berasal dari sel yang sudah ada sebelumya yang telah berkembang biak, seperti yang dijelaskan oleh Rudolf Virchow pada tahun 1858 ketika menyatakan omnis cellula e cellula (semua sel berasal dari sel)
Penelitian Virchow membantu membangun konsep patologi seluler, gagasan bahwa semua penyakit disebabkan oleh perubahan sel normal. Virchow berpendapat bahwa kehidupan hanyalah hasil dari proses aktivitas seluler. Pada tahun 1855, Virchow mengembangkan lebih lanjut tentang ide nya dengan menerbitkan pepatah yang terkenal “omnis cellula e cellula” yang menjadi dasar teori sel. Teori Virchow menyatakan bahwa sama seperti hewan tidak dapat muncul tanpa hewan yang sudah ada sebelumnya, sel tidak dapat muncul tanpa sel yang ada sebelumnya. Gagasan bahwa sel -sel baru muncul dari sel yang sudah ada sebelumnya, baik di jaringan yang sakit maupun sehat bukan gagasan asli dari Virchow.
Walter Flemming merupakan ahli anatomi Jerman, dan pendiri ilmu sitogenetika.Flemming merupakan orang pertama yang mengamati dan menggambarkan secara sistematis perilaku kromosom di dalam inti sel selama pembelahan sel (mitosis). Setelah menjadi dokter militer selama perang Prancis-Jerman, Flemming memperoleh posisi penting di Universitas Praha dan Universitas Kiel. Flemming merupakan pelopor penggunaan pewarna aniline yang baru ditemukannya untuk memvisualisasikan struktur sel, dan pada tahun 1879 dia menyatakan bahwa beberapa pewarna tertentu mampu mewarnai benang spindel di dalam nukleus. Melalui penemuannya ini, Flemming menjelaskan serangkaian peristiwa penting yang terjadi di nukleus selama pembelahan sel. Dia menunjukkan bahwa benang spindel memendek dan tampaknya membelah secara longitudinal menjadi dua bagian dan menuju kearah yang berlawanan. Seluruh hasil risetnya terkait mitosis ditulis dalam buku bersejarah yang berjudul “Zell-substanz, Kern und Zelltheilung” atau yang berarti “Cell-Substance, Nucleus, and Cell-Division”. Tetapi penemuan Flemming terhadap peran mitosis dalam hereditas tidak sepenuhnya dihargai, hingga pengakuan prinsip-prinsip hereditas Gregor Mendel dibuka kembali 20 tahun kemudian.
Wilhelm August Oscar Hertwig berkontribusi pada ilmu embriologi melalui studinya tentang sel yang dalam penemuannya menyatakan bahwa satu spermatozoa diperlukan untuk membuahi sel telur. Hertwig awalnya mengabdikan dirinya untuk mempelajari perkembangan morfologis, topik yang ditulisnya dalam disertasi doktoralnya di Bonn pada tahun 1872. Nanum ia beralih mempelajari sifat proses pembuahan, setelah membaca organologische studien karya Leopold Auebach. Pada masa itu, pandangan utama terkait embriologi adalah bahwa spermatozoa melakukan kontak dengan telur dan merangsang perkembangan melalui transmisi getaran mekanis, atau spermatozoa menembus telur dan bercampur dengan komponen kimianya. Hertwig mampu menemukan proses pembuahan itu pada Bulu Babi (Sea urchin), dimana ia menemukan bahwa hanya satu spermatozoa yang memasuki sel telur, dan sel telur tersebut akan membentuk membran viteline yang menghalangi masuknya spermatozoa lainnya.
Max Schultze belajar kedokteran di Greifswald dan Berlin serta diangkat sebagai profesor luar biasa di Halle pada 1854 dan lima tahun kemudian diangkat menjadi profesor biasa di bidang anatomi dan histologi oleh Direktur Institut Anatomi di Bonn. Max Schultze dikenal karena karyanya pada teori tentang sel. Dengan menggabungkan teori Felix Dujardin dan kosep Sarcode pada binatang dengan Hugo Von Mohl dengan protoplasma pada sayuran. Schultze menegaskan bahwa protoplasma merupakan dasar-dasar fisik kehidupan. Protoplasma bukan hanya bagian struktural sel, tetapi juga merupakan bagian penting sel sebagai tempat berlangsung reaksi-reaksi kimia kehidupan.Protoplasma juga merupakan tempat terjadinya proses hidup.
Milestone Perkembangan Biologi Sel
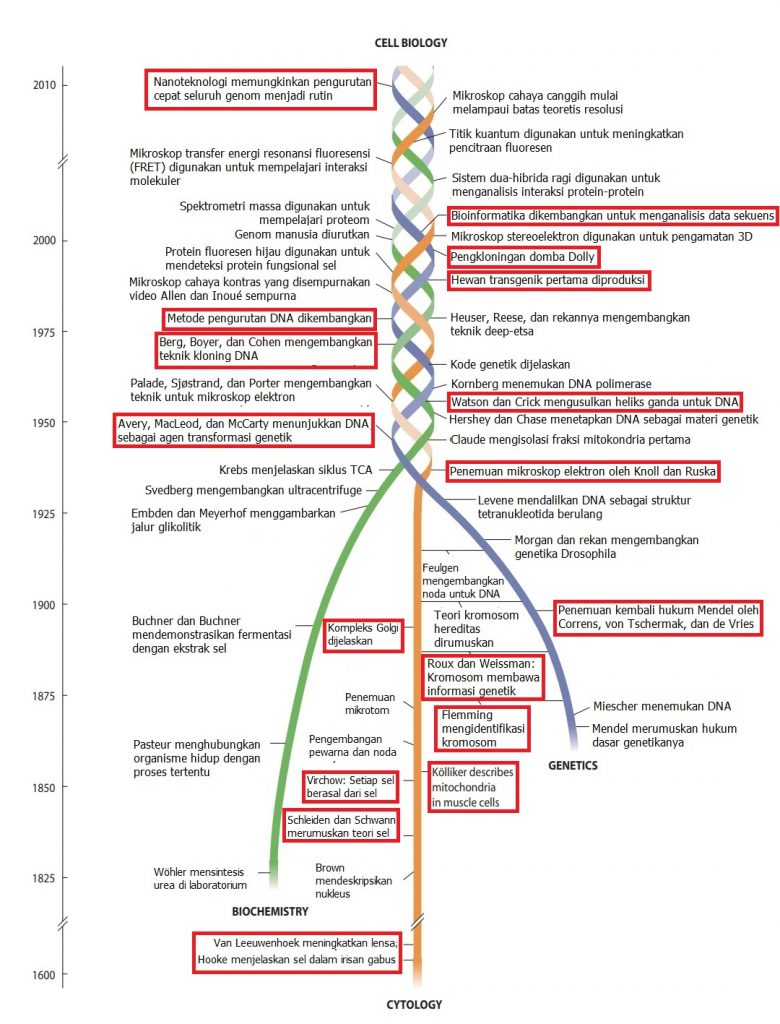
Millestone perkembangan ilmu terkait biologi sel tidak bisa dilepaskan dari perkembangan ilmu genetika dan biokimia. Perkembangan ilmu tentang sel diawali dengan penemuan oleh Robert Hooke dan Anthony van Leeuwenhoek tentang sel, sedangkan ilmu genetika diawali dengan formulasi hukum pewarisan sifat yang ditemukan oleh Mendel. Perkembangan ilmu biokimia diawali dengan penemuan Wohler tentang sintesis urea di laboratorium. Setelah tahun 1950, temuan terkait double helix DNA oleh Watson dan Crick merupakan kombinasi pengetahuan sel, biokimia, dan genetika. Hingga saat ini, perkembangan ilmu terkait dengan ini telah berkembang jauh hingga kajian bioinformatika dan nanoteknologi.
Perkembangan Teknologi Mikroskop dalam Menganalisis Sel
Mikroskop merupakan salah satu cara dan cara yang paling efektif dalam mempelajari. Perkembangan terkait dengan penemuan tentang sel sangat dipengaruhi oleh perkembangan penemuan terkait mikroskop. Cerita tentang awal perkembangan sel oleh Robert Hooke dan Anthony van Leeuwenhoek menunjukkan bahwa mikroskop sangat berperan penting dalam penemuan terkait sel.
Antonie van Leeuwenhoek terkenal sebagai bapak Mikrobiologi karena merupakan orang pertama yang mampu menciptakan mikroskop yang cukup kuat untuk melihat mikroba. Meskipun mikroskop van Leeuwenhoek merupakan mikroskop sederhana, namaun sangat kuat dan memberikan resolusi yang lebih baik daripada mikroskop majemuk pada zamannya. Meskipun terkenal dalam mengembangkan teleskop, Galileo Galilei juga merupakan salah satu pelopor pengembang mikroskop. Jadi siapa sebenarnya orang yang pertama kali mendesain mikroskop? Jawabannya adalah Hans dan Zaccharias Janssen, seorang pembuat kacamata yang mungkin juga telah menemukan teleskop, mikroskop sederhana, dan mikroskop majemuk pada akhir tahun 1500-an atau awal 1600-an. Sayangnya sedikit informasi tentang Janssens, sedangkan pekerjaannya tidak pernah dipublikasikan.
Mikroskop milik Leeuwenhook ini masih terdapat kekurangan pada lensa cembung. Cara menggunakan mikroskop lab milik Leeuwenhook ini juga tidak sesimpel seperti yang Anda temui hari ini. Sehingga berbagai peneliti lainnya melakukan perkembangan dengan menambah lensa pendukung yang sekarang dikenal dengan lensa obyektif. Mikroskop terus mengalami perkembangan, seperti ada penambahan cermin sebagai sumber pencahayaan, kondensor, dan sebagainya. Hingga mikroskop modern mampu memperbesar obyek hingga 1.000 kali. Pada 1920 saat ditemukan adanya elektron, hingga pada 1932 berkembanglah mikroskop elektron yang bisa memperbesar ukuran benda hingga dua juta kali lipat. Anda pun kini bisa memahami dengan mudah bagaimana cara menggunakan mikroskop lab, tidak seperti dulu lagi dengan berbagai macam kekurangan.
Saat ini, kebayakan laboratorium telah memiliki minimal mikroskop cahaya, walaupun ternyata banyak jenis mikroskop yang telah dikembangkan, dan sangat tergantung dengan kebutuhan dari pengamatan yang dilakukan. Mulai dari mikroskop cahaya yang hanya untuk melihat tampilan sederhana dari objek, hingga mikroskop elektron dengan tampilan tiga dimensi dari objek yang sangat kecil, misalnya organel sel.
Mikroskop cahaya menghasilkan dua bidang pengamatan, bergantung apda kebutuhan peneliti. Bidang yang dimaksud adalah fase kontras dan bidang terang. Mikroskop fase kontras mampu mengubah perbedaan indeks bisa menjadi perbedaan kecerahan, sehingga keberadaan sedikit udara dalam spesimen dapat diminimalisir pandangannya, sedangkan pada fase terang akan terlihat jelas, karena gelembung udara tersebut memiliki indeks bisa yang berbeda dengan medium sekitarnya. Keuntungan mikroskop tampilan bidang terang adalah optik tidak mengubah warna struktur yang diamati, penggunaan warna terkadang membuat struktur tertentu terlihat jelas, umumnya harganya lebih murah dibandingkan dengan mikroskop denganf ase kontras serta mikroskop ini membutuhkan sedikit penyesuaian sebelum seseorang dapat mengamati spesimen. Keuntungan mikroskop fase kontras yaitu memungkinkan visualisasi struktur tertentu yang sebaliknya tidak terlihat, termasuk organel tertentu yang tidak dapat diamati pada bidang terang, dan terkadang gambar fase kontras secara subyektif terlihat lebih baik daripada gambar bidang terang karena detail dari objek dapat terlihat.
Mikroskop fluoresensi hampir sama dengan mikroskop cahaya konvensinal. Mikroskop fluoresensi di sisilain menggunakan sumber cahaya dengan intensitas cahaya yang jauh lebih tinggi dan kuat seperti pada spesies fluoresens dalam sampel bunga atau jamur. Mikroskop fluoresensi sering digunakan untuk menggambarkan fitur spesifik dari spesimen kecil seperti mikroba. Mikroskop ini juga digunakan untuk meningkatkan fitur tiga dimensi secara visual dalam skala kecil. Hal ini dapat diperoleh dengan menempelkan pewarna fluoresensi ke anti-bodi yang pada akhirnya menempel pada fitur yang ditargetkan. Ketika cahaya dipantulkan fan fluoresensi background difilter dalam jenis mikroskop ini, bagian yang ditergitkan dari sampel dapat dicitrakan dengan jelas. Ini memberikan para peneliti untuk mampu memvisualisasikan organel yang diinginkan atau fitur permukaan unit dari sampel yang diinginkan. Mikroskop fluoresensi confocal paling sering digunakan untuk menonjolkan sifat 3D sampel. Ini dihasilkan menggunakan sumber cahaya yang kuat, seperti laser yang dapat difokuskan dengan tepat. Pemfokusan ini dilakukan berulang kali sepanjang satu tingkat spesimen.
Mikroskop confocal secara luas digunakan untuk menyelesaikan struktur rinci objek tertentu di dalam sel. Mirip dengan mikroskop fluoresensi, berbagai komponen sel hidup dan tetap atau bagian jaringan dapat diberi label khusus menggunakan imunofluoresensi, misalnya dan kemudian divisualisasikan dalam resolusi yang tinggi. Sebagai ciri khas, mikroskop confocal memungkinkan pembuatan gambar yang tajam dari fokus yang tepat, tanpa cahaya fluoresensi yang mengganggu dari background atau bagian lain dari spesimen. Oleh karena itu, struktur dalam objek yang lebih tebat dapat dengan mudah divisualisasikan menggunakan mikroskop confocal. Selanjutnya dengan menumpuk beberapa gambar dari bidang optik yang berbeda, struktur 3D dapat dianalisis. Mikroskop confocal memiliki kedalaman penetrasi sampel yang terbatas, objek yang lebih tebal dicitrakan secara optimal menggunakan mikroskop tipe confocal LSFM atau mikroskop dua foton.
Mikroskop elektron adalah mikroskop yang menggunakan berkas elektron yang diakselerasi sebagai sumber penerangan. Mikroskop khusus jenis ini memiliki resolusi gambar yang tinggi, mampu memperbesar objek dalam nanometer yang dibentuk menggunakan elektron terkontrol dalam luar hampa dan ditangkap pada layar yang berpendar. Mikroskop elektron pertama dibangun pertama kali oleh teknisi Jerman bernama Ernst Ruska pada tahun 1931.
Ketika sebuah film diputar di teater, cahaya ditransmisikan melalui gambar pada film. Saat berkas cahaya melewati, itu dimodifikasi oleh gambar dan isi film kemudian ditampilkan. TEM bekerja dengan cara yang sama tetapi dengan elektron, melewati, atau mentransmisikan, sampel ultra tipis ke detektor di bawah. TEM memungkinkan Anda untuk mengamati detail sekecil atom individu, memberikan tingkat informasi struktural yang belum pernah terjadi sebelumnya pada resolusi setinggi mungkin. Saat melewati objek, itu juga dapat memberi Anda informasi tentang struktur internal, yang tidak dapat diberikan oleh SEM. Namun, TEM terbatas pada sampel yang cukup tipis untuk membiarkan elektron melewatinya. Proses penipisan ini secara teknis menantang dan membutuhkan alat tambahan untuk melakukannya.
Bayangkan Anda berada di ruangan gelap dengan senter cahaya yang lemah. Untuk menjelajahi lingkungan Anda, Anda dapat menggeser cahaya melintasi ruangan, seperti seseorang yang membaca buku: kiri ke kanan dan atas ke bawah. SEM berfungsi sama, menyapu berkas elektron melintasi sampel dan merekam elektron yang memantul kembali. Teknik ini memungkinkan Anda untuk melihat permukaan hampir semua sampel, dari logam industri hingga sampel geologis hingga spesimen biologis seperti spora, serangga, dan sel. Meskipun SEM tidak dapat melihat fitur dengan tingkat detail yang sama seperti TEM, SEM jauh lebih cepat, tidak terlalu membatasi, dan terkadang dapat dilakukan dengan persiapan sampel yang terbatas atau tanpa persiapan.
Ukuran Sel
Sel terbagi dalam berbagai ukuran dan bentuk. Beberapa sel bakteri terkecil misalnya, hanya berdiameter sekitar 0,2–0,3 mm. Sangat kecil sehingga sekitar 50.000 sel semacam itu bisa muat berdampingan di ibu jari Anda. Di sisi lain terdapat sel-sel saraf yang sangat panjang hingga mencapai ukuran satu meter atau lebih. Sel saraf sepanjang leher atau kaki jerapah adalah contoh nya. Begitu juga telur burung yang merupakan sel tunggal yang sangat besar, meskipun sebagian besar volume internalnya ditempati oleh kuning telur yang merupakan makanan embrio yang sedang berkembang. Meskipun begitu, sebagian besar sel berada dalam kisaran ukuran yang kecil. Misalnya sel bakteri dan archaeal yang biasanya berdiameter sekitar 1-5 mm, sedangkan sebagian besar sel tumbuhan dan hewan tingkat tinggi memiliki dimensi dalam kisaran 10-100 mm. Mengapa sel sangat kecil? Berbagai faktor diketahui mempengaruhi ukuran sel, namun 3 faktor diantaranya yang paling berpengaruh adalah (1) kebutuhan akan rasio luas permukaan/volume yang memadai, (2) kecepatan difusi molekul, dan (3) kebutuhan untuk mempertahankan konsentrasi dari molekul spesifik seperti zat dan enzim yang terlibat dalam proses seluler. Sekarang mari kita bahas masing-masing dari ketiga faktor tersebut.
Luas Permukaan/Rasio Volume
Dalam kebanyakan kasus, batasan utama pada ukuran sel ditentukan oleh kebutuhan untuk mempertahankan rasio luas permukaan/volume. Luas permukaan sangat penting karena pada permukaan sel terjadi pertukaran yang diperlukan antara sel dan lingkungannya. Volume internal sel menentukan jumlah nutrisi yang harus diimpor dan jumlah produk limbah yang harus dikeluarkan. Luas permukaan secara efektif mewakili jumlah membran sel yang tersedia untuk penyerapan dan ekskresi tersebut. Masalah dalam mempertahankan luas permukaan muncul karena volume sel meningkat dengan pangkat tiga dari panjang atau diameter sel, sedangkan luas permukaannya meningkat hanya dengan kuadrat. Dengan demikian, sel besar memiliki rasio luas permukaan terhadap volume yang lebih rendah daripada sel kecil. Perbandingan ini mengilustrasikan kendala utama pada ukuran sel yaitu “seiring dengan bertambahnya ukuran sel, luas permukaannya tidak sebanding dengan volumenya, dan pertukaran zat yang diperlukan antara sel dan lingkungan sekitarnya menjadi semakin bermasalah.” Oleh karena itu, ukuran sel dapat meningkat hanya selama luas permukaan membran masih cukup untuk masuk dan keluarnya bahan dari sel.
Beberapa sel, terutama yang berperan dalam penyerapan, memiliki karakteristik yang memaksimalkan luas permukaannya. Luas permukaan efektif paling sering ditingkatkan oleh lipatan ke dalam atau penonjolan membran sel ke luar. Sel-sel yang melapisi usus kecil misalnya, mengandung banyak tonjolan yang disebut mikrovili yang sangat meningkatkan luas permukaan membran dan menigkatkan kapasitas penyerapan nutrisi.
Tingkat Difusi Molekul
Ukuran sel juga berpengaruh terhadap kecepatan pergerakan molekul di dalam sel dalam menjalankan aktivitas seluler tertentu. Banyak molekul bergerak melalui difusi, yang merupakan pergerakan bebas suatu zat dari daerah dengan konsentrasi tinggi ke daerah dengan konsentrasi rendah. Kecepatan pergerakan molekul melaui difusi dipengaruhi oleh ukuran molekul dan laju difusi menurun seiring dengan bertambahnya ukuran molekul. Penelitian terbaru menunjukkan bahwa banyak sel eukariotik dapat melewati batasan ini dengan secara aktif mengangkut ion, makromolekul, dan bahan lain melalui sitoplasma menggunakan protein pembawa khusus. Beberapa sel organisme yang lebih tinggi mengatasi keterbatasan ini dengan aliran sitoplasma (juga disebut siklosis dalam sel tumbuhan), suatu proses yang melibatkan gerakan aktif dan pencampuran isi sitoplasma. Sel lain memindahkan molekul tertentu melalui menggunakan vesikel yang diangkut sepanjang mikrotubulus. Dengan tidak adanya mekanisme ini, bagaimanapun, ukuran sel dibatasi oleh tingkat difusi molekul yang dikandungnya.
Kebutuhan Konsentrasi Reaktan dan Katalis yang Memadai
Keterbatasan ketiga pada ukuran sel adalah kebutuhan untuk mempertahankan konsentrasi yang memadai dari senyawa esensial dan enzim yang dibutuhkan untuk berbagai proses yang harus dilakukan sel. Agar reaksi kimia terjadi dalam sel, reaktan yang sesuai harus bertabrakan dan berikatan dengan enzim tertentu. Frekuensi tumbukan tersebut akan sangat meningkat dengan konsentrasi yang lebih tinggi dari reaktan dan enzim. Untuk mempertahankan konsentrasi molekul tertentu, jumlah molekul harus meningkat secara proporsional dengan volume sel. Setiap kali masing-masing dari tiga dimensi sel berlipat ganda, ada peningkatan delapan kali lipat dalam volume sel, dan dengan demikian diperlukan delapan kali lebih banyak molekul untuk mempertahankan konsentrasi aslinya. Dengan tidak adanya mekanisme pemusatan, ini jelas membebani kemampuan sintetik sel.
Sel ada dalam berbagai bentuk dan ukuran, dari sel jamur berfilamen hingga bakteri Treponema berbentuk spiral hingga sel-sel berbentuk berbeda dari sistem darah manusia. Sel lain memiliki bentuk yang jauh lebih eksotis, seperti radiolaria dan protozoa. Bahkan dua gamet bersel tunggal manusia, telur dan sperma, sangat berbeda dalam ukuran dan bentuk. Seringkali, apresiasi terhadap bentuk dan struktur sel memberikan petunjuk tentang fungsinya. Misalnya, luas permukaan mikrovili yang besar pada sel usus kita membantu memaksimalkan penyerapan nutrisi, penebalan spiral di dinding sel jaringan xilem tanaman memberi kekuatan pada pembuluh penghantar air di kayu, dan sel-sel neuron manusia yang sangat bercabang memungkinkannya berinteraksi dengan banyak neuron lain.
Berbagai Jenis Sel
Ada banyak perbedaan antara bakteri, archaea, dan eukariota dalam hal struktural, biokimia, dan genetik yang penting di antara kelompok-kelompok ini. Beberapa dari perbedaan ini diringkas dan dapat dilihat pada Tabel 1 di bawah. Untuk saat ini, penting untuk disadari bahwa meskipun ada beberapa kesamaan karakteristik di antara sel-sel dari masing-masing tiga domain ini, setiap jenis sel memiliki seperangkat sifat pembeda yang unik.
Kehadiran Inti Terikat Membran
Perbedaan struktural secara tradisional telah dibuat antara eukariota dan prokariota (bakteri dan archaea) dan tercermin dalam nomenklatur itu sendiri. Namun, saat kita mempelajari lebih lanjut tentang detail seluler dari tiga domain organisme ini, perbedaan ini menjadi kurang penting dibandingkan aspek lain yang dilihat dari struktur dan fungsi. Sel eukariotik memiliki inti sejati yang dibatasi membran, sedangkan sel prokariotik tidak. Alih-alih diselimuti oleh membran, informasi genetik dari sel bakteri atau archaeal dilipat menjadi struktur kompak yang dikenal sebagai nukleoid, yang melekat pada membran sel di wilayah tertentu dari sitoplasma. Di sisi lain dalam sel eukariotik, sebagian besar informasi genetik terlokalisasi pada nukleus, yang dikelilingi bukan oleh membran tunggal tetapi oleh selubung nukleus yang terdiri dari dua membran. Nukleus juga mencakup nukleolus, yang merupakan tempat sintesis RNA ribosom dan perakitan subunit ribosom, dan mengandung kromosom pembawa DNA, yang tersebar sebagai kromatin di seluruh nukleoplasma setengah cair yang mengisi volume internal nukleus.
Kehadiran Membran Internal untuk Memisahkan Fungsi
Seperti yang dijelaskan sebelumnya, sel bakteri (dan archaeal) umumnya tidak mengandung membran internal dan sebagian besar fungsi seluler terjadi baik di sitoplasma atau pada membran plasma. Namun, ada sekelompok bakteri fotosintetik yang dikenal sebagai Cyanobacteria yang memiliki membran internal yang luas di mana reaksi fotosintesis dilakukan. Juga, beberapa bakteri memiliki struktur terikat membran yang menyerupai organel, sementara yang lain memiliki kompartemen berlapis protein yang berfungsi sebagai organel dengan mengisolasi kelompok enzim yang terlibat dalam jalur metabolisme tertentu. Sebaliknya, hampir semua sel eukariotik menggunakan membran internal secara ekstensif untuk mengkotak-kotakkan fungsi tertentu , dan mereka sering memiliki banyak organel.
Contoh sistem membran internal pada sel eukariotik termasuk retikulum endoplasma, kompleks Golgi, dan membran yang mengelilingi dan membatasi organel seperti mitokondria, kloroplas, lisosom, dan peroksisom, serta berbagai jenis vakuola dan vesikel. Masing-masing organel ini dikelilingi oleh membran karakteristiknya sendiri (atau sepasang membran) yang mungkin mirip dengan membran lain dalam struktur dasar tetapi dapat memiliki komposisi kimia yang khas. Terlokalisasi dalam setiap organel tersebut adalah mesin molekuler yang diperlukan untuk melaksanakan fungsi seluler tertentu yang strukturnya khusus.
Sitoskeleton
Sitoskeleton juga ditemukan dalam sitoplasma sel eukariotik beberapa nonmembran, struktur protein yang terlibat dalam kontraksi seluler, motilitas, dan pembentukan serta dukungan arsitektur seluler. Termasuk juga mikrotubulus yang ditemukan di silia dan flagela dari banyak jenis sel, mikrofilamen yang ditemukan di fibril otot dan struktur lain yang terlibat dalam motilitas, dan filamen perantara, yang sangat menonjol dalam sel yang mengalami stress atau tekanan. Mikrotubulus, mikrofilamen, dan filamen intermediet adalah komponen kunci dari sitoskeleton, yang memberikan struktur dan elastisitas pada hampir semua sel eukariotik. Selain itu, sitoskeleton dapat menyediakan perancah untuk intraseluler transportasi vesikel ke tempat-tempat di dalam sel di mana isinya dibutuhkan. Baru-baru ini, protein yang mirip dengan protein sitoskeleton telah ditemukan pada bakteri dan tampaknya memiliki peran dalam mempertahankan bentuk sel.
Eksositosis dan Endositosis
Fitur lebih lanjut dari sel eukariotik adalah kemampuannya untuk bertukar bahan antara kompartemen yang dibatasi membran di dalam sel dan bagian luar sel. Pertukaran ini dimungkinkan karena eksositosis dan endositosis, proses yang melibatkan peristiwa fusi membran yang unik untuk sel eukariotik. Pada endositosis, bagian dari membran plasma berinvaginasi dan terjepit untuk membentuk vesikel sitoplasma berbatas membran yang mengandung zat yang sebelumnya berada di luar sel. Eksositosis pada dasarnya adalah kebalikan dari proses ini: Vesikel yang dibatasi membran di dalam sel menyatu dengan membran plasma dan melepaskan isinya ke luar sel.
Perbedaan Sel Prokariotik dan Eukariotik
Tabel 1. Perbedaan sel prokarotik dan eukariotik
| Perbedaan | Prokariotik | Eukariotik |
| Ukuran Sel | 1-10 um | 10-100 um |
| Metabolisme | Anaerobik dan Aerobik | Aerobik |
| Cara Hidup | Autotrof | Autotrof dan heterotrof |
| Organela | Tidak ada | Ada berbagai jenis seperti nukleus, mitokondria, retikulum endoplasma, kompleks golgi, dan kloroplas |
| DNA | Terletak dalam sitoplasma, struktur sirkuler | Terletak pada nukleoplasma, struktur linier (mirip benang halus yang sangat panjang) dan segmental |
| Struktur Gen | Linier | Morsel (Segmental) |
| Nukleolus | Tidak ada | Ada |
| Jumlah kromosom | Satu | Lebih dari satu |
| RNA dan Protein | RNA dan protein disintesis pada ruang yang sama | RNA disintesis dalam nukleus, protein dan sitoplasma |
| Sitoplasma | Tanpa sitoseklet, tidak ada gerakan sitoplasmik, proses endositosis maupun eksositosis | Memiliki sitoskelet, terjadi gerakan sitoplasmik, proses endositosis dan eksositosis |
| Motilitas | Dengan flagella dan silia | Mikrotubul dan tubulin, aktin dan miosin |
| Pembelahan sel | Amitosis, kromatin ditarik dengan jaran melekat pada selaput plasma | Mitosis, kromosom dipisahkan oleh aparatus mitosis yang terdiri dari filamen sitoskeletik |
| Ribosom | 70 S dengan 3 rRNA | 80 S dengan 4 rRNA |
| Seksualitas | Konjugasi | Meiosis |
| Organisasi seluler | Sebagian unisel | Sebagian besar multisel dengan diferensiasi menjadi berbagai jenis sel |
Organisasi DNA
Perbedaan lain antara bakteri, archaea, dan eukariota adalah jumlah dan organisasi materi genetik. DNA bakteri biasanya hadir dalam sel sebagai molekul melingkar yang terkait dengan protein yang relatif sedikit.Di sisi lain, DNA eukariotik ada di dalam sel sebagai beberapa molekul linier yang dikomplekskan dengan sejumlah besar protein yang dikenal sebagai histon. DNA archaeal biasanya melingkar dan dikomplekskan dengan protein dalam jumlah sedang yang menyerupai protein histon eukariotik
Molekul DNA sirkular dari sel bakteri atau archaeal jauh lebih panjang daripada sel itu sendiri. Oleh karena itu harus dilipat dan dikemas bersama-sama erat untuk masuk ke dalam sel. Misalnya, bakteri usus biasa Escherichia coli hanya memiliki panjang sekitar satu atau dua mikrometer, namun memiliki molekul DNA melingkar dengan keliling sekitar 1300 mm. Jelas, banyak lipatan dan pengemasan diperlukan untuk memasukkan DNA sebanyak itu ke dalam sel sekecil itu. Dengan analogi, kira-kira setara dengan mengemas sekitar 60 kaki (18 m) benang yang sangat tipis ke dalam bidal biasa.
Tetapi jika DNA tampaknya menimbulkan masalah pengemasan untuk sel prokariotik, pertimbangkan kasus sel eukariotik. Kebanyakan sel eukariotik memiliki setidaknya 1000 kali lebih banyak DNA seperti yang dimiliki E. coli tetapi hanya mengkodekan 5-10 kali lebih banyak protein. Karena para ilmuwan tidak mengetahui fungsi dari kelebihan, DNA noncoding itu sering disebut sebagai “DNA sampah.” Namun tampaknya banyak dari DNA noncoding ini mungkin memiliki fungsi penting selain pengkodean protein seluler. Beberapa di antaranya terlibat dalam produksi mRNA pengatur, beberapa terlibat dalam generasi keanekaragaman spesies selama evolusi, dan beberapa mengandung urutan berulang yang tampaknya berfungsi sebagai situs pengikatan untuk protein pengatur. Sebuah studi baru-baru ini menunjukkan bahwa hingga 33% dari situs pengikatan untuk faktor transkripsi dengan peran dalam perkembangan kanker ditemukan di wilayah DNA berulang ini. Jadi, kita harus berhati-hati untuk tidak berasumsi bahwa tidak ada fungsi untuk komponen seluler tertentu hanya karena saat ini kita tidak memahami perannya di dalam sel.
Apa pun fungsi DNA dalam jumlah besar seperti itu, masalah pengemasan semua bahan genetik ini diselesaikan secara universal di antara eukariota dengan organisasi DNA ke dalam struktur kompleks yang disebut kromosom, yang mengandung setidaknya protein histon sebanyak DNA. Sebagai kromosom DNA sel eukariotik dikemas, dipisahkan selama pembelahan sel, dan ditransmisikan ke sel anak.
Pemisahan Informasi Genetik
Kontras lebih lanjut antara prokariota dan eukariota adalah cara mereka mengalokasikan informasi genetik ke sel anak pada saat pembelahan. Sel bakteri dan archaeal hanya mereplikasi DNA mereka dan membelah dengan proses yang relatif sederhana yang disebut pembelahan biner, dengan satu molekul DNA yang direplikasi dan setengah dari sitoplasma pergi ke setiap sel anak. Setelah replikasi DNA dalam sel eukariotik, kromosom didistribusikan secara merata ke sel anak melalui proses mitosis dan meiosis yang lebih kompleks, diikuti oleh sitokinesis, pembelahan sitoplasma.
Evolusi Sel
Mitokondria dan kloroplas merupakan organel eukariotik, namun terdapat teori yang membahas mengenai asal usul evolusi organel ini. Baik mitokondria maupun kloroplas mengandung DNA dan ribosom mereka sendiri, yang memungkinkan mereka melakukan sintesis RNA dan protein (walaupun sebagian besar protein yang ada dalam organel ini sebenarnya dikodekan oleh gen nuclear). Ketika ahli biologi molekuler mempelajari asam nukleat dan sintesis protein dalam organel ini, mereka dikejutkan oleh banyak kesamaan antara proses ini di mitokondria dan kloroplas dengan proses dalam sel bakteri. Keduanya memiliki molekul DNA melingkar tanpa histon terkait, dan keduanya menunjukkan kesamaan dalam urutan rRNA, ukuran ribosom, kepekaan terhadap inhibitor RNA dan sintesis protein, dan jenis faktor protein yang digunakan dalam sintesis protein. Selain fitur molekuler ini, mitokondria dan kloroplas menyerupai sel bakteri dalam ukuran dan bentuk, dan mereka memiliki membran ganda di mana membran dalam memiliki lipid tipe bakteri.
Kesamaan ini mengarah pada teori endosimbion untuk asal usul evolusi mitokondria dan kloroplas. Teori ini, yang dijelaskan lebih lengkap dalam Kotak 11A, halaman 298–299, mengusulkan bahwa kedua organel ini berasal dari prokariota yang masuk dan membentuk hubungan simbiosis di dalam sitoplasma organisme bersel tunggal purba yang disebut protoeukariota. Teori endosimbion mengusulkan bahwa protoeukariota menelan bakteri dan cyanobacteria melalui proses yang dikenal sebagai fagositosis (“pemakan sel”). Setelah fagositosis, sel-sel ini tidak dicerna; sebaliknya, mereka mengambil tempat tinggal di sitoplasma dan akhirnya berkembang menjadi mitokondria dan kloroplas, masing-masing.
Perdebatan tentang asal usul evolusi mitokondria dan kloroplas memiliki sejarah panjang. Pada awal tahun 1883, Andreas F. W. Schimper menyarankan bahwa kloroplas muncul dari hubungan simbiosis antara bakteri fotosintetik dan sel nonfotosintetik. Pada pertengahan 1920-an, peneliti lain telah memperluas ide Schimper dengan mengusulkan asal simbiosis mitokondria. Ide-ide tersebut mendapat ejekan dan pengabaian selama beberapa dekade, namun, sampai tahun 1960-an, ketika ditemukan bahwa mitokondria dan kloroplas mengandung DNA mereka sendiri. Penelitian lebih lanjut mengungkapkan bahwa mitokondria dan kloroplas adalah organel semiotonom, organel yang dapat membelah sendiri dan tidak hanya mengandung DNA tetapi juga mRNA, tRNA, dan ribosom.
Kita sekarang menyadari bahwa proses DNA, RNA, dan sintesis protein di mitokondria dan kloroplas lebih mirip dengan proses analog di sel bakteri daripada di sel eukariotik. Hal ini menyebabkan para ahli biologi merumuskan teori endosimbion. Teori ini, dikembangkan sepenuhnya oleh Lynn Margulis, mengusulkan bahwa mitokondria dan kloroplas berevolusi dari bakteri purba yang membentuk hubungan simbiosis (hubungan yang saling menguntungkan) dengan sel berinti primitif 1 hingga 2 miliar tahun yang lalu..
Asumsi awal dari teori endosimbion adalah bahwa tidak adanya molekul oksigen di atmosfer primitif Bumi membatasi sel-sel awal pada mekanisme anaerobik untuk memperoleh energi. Beberapa sel anaerobik kemudian mengembangkan pigmen yang mampu mengubah energi cahaya menjadi energi kimia, memungkinkan mereka menggunakan sinar matahari sebagai sumber energi. Beberapa organisme fotosintesis ini mengembangkan mekanisme untuk menggunakan air sebagai donor elektron. Akibatnya, oksigen dilepaskan sebagai produk sampingan dan komposisi atmosfer bumi berubah secara dramatis.
Ketika oksigen terakumulasi di atmosfer bumi, beberapa bakteri anaerob berevolusi menjadi organisme aerobik dengan mengembangkan transpor elektron yang bergantung pada oksigen dan jalur fosforilasi oksidatif. Teori endosimbion menunjukkan bahwa nenek moyang sel eukariotik (disebut protoeukariota) mengembangkan setidaknya satu fitur penting yang membedakannya dari sel primitif lainnya: kemampuan untuk menelan nutrisi dan partikel dari lingkungan melalui fagositosis. Karakteristik ini memungkinkan protoeukariota untuk membangun hubungan endosimbiosis dengan bakteri primitif. Bagaimana sel-sel primitif ini memperoleh nukleus yang terikat membran masih belum jelas.
Mitokondria Ternyata Berevolusi dari Bakteri Ungu Kuno
Langkah pertama menuju evolusi mitokondria mungkin terjadi ketika protoeukariota anaerobik yang bergantung pada glikolisis untuk energi menelan bakteri aerob yang lebih kecil dengan fagositosis. Bakteri aerob yang tertelan mampu melakukan respirasi aerob dan akan menyediakan sejumlah besar energi yang berguna untuk sel inang anaerobik. Pada gilirannya, sel inang memberikan perlindungan dan nutrisi kepada bakteri yang berada di sitoplasmanya. Dengan demikian, bakteri yang tertelan dan protoeukariota membentuk hubungan simbiosis yang saling menguntungkan. Secara bertahap, selama ratusan juta tahun, bakteri kehilangan fungsi yang tidak penting dalam lingkungan sitoplasma baru mereka dan berkembang menjadi mitokondria. Ketika urutan dasar RNA ribosom mitokondria (rRNA) dibandingkan dengan berbagai rRNA bakteri, kecocokan paling dekat terjadi di antara bakteri ungu, menunjukkan bahwa nenek moyang mitokondria yang tertelan adalah anggota kuno dari kelompok ini.
Kloroplas Ternyata Berevolusi dari Cyanobacteria Kuno
Menurut teori endosimbion, langkah pertama menuju evolusi kloroplas terjadi ketika anggota subkelompok eukariota awal, yang sudah dilengkapi dengan bakteri aerob atau mitokondria primitif, menelan sel fotosintesis primitif. Seperti yang baru saja dijelaskan untuk evolusi mitokondria, organisme yang tertelan mungkin menyediakan energi untuk sel inang sebagai ganti tempat berlindung dan nutrisi. Sel-sel fotosintesis secara bertahap kehilangan fungsi yang tidak penting dalam lingkungan baru mereka dan berkembang menjadi komponen integral dari inang eukariotik. Ketika urutan dasar rRNA kloroplas dibandingkan dengan berbagai rRNA bakteri, kecocokan paling dekat terjadi di antara cyanobacteria, menunjukkan bahwa nenek moyang kloroplas adalah anggota kuno dari kelompok ini.
Teori endosimbion terutama didasarkan pada kesamaan biokimia yang diamati di antara mitokondria, kloroplas, dan bakteri, tetapi dukungan juga diberikan oleh hubungan simbiosis kontemporer yang menyerupai apa yang mungkin terjadi di masa lalu. Alga, dinoflagellata, diatom, dan bakteri fotosintetik hidup sebagai endosimbion dalam sitoplasma sel yang terdapat pada lebih dari 150 jenis protista dan invertebrata yang ada. Dinding sel organisme yang tertelan seringkali tidak ada lagi, dan dalam beberapa kasus struktur sel bahkan semakin berkurang, dengan hanya kloroplas dari endosimbion yang tersisa.
Contoh mencolok dari endosimbiosis terjadi pada siput laut tertentu dan moluska terkait, di mana sel-sel yang melapisi saluran pencernaan hewan mengandung kloroplas yang dapat diidentifikasi dengan jelas. Kloroplas ini, yang berasal dari ganggang hijau yang dimakan moluska, terus melakukan fotosintesis lama setelah dimasukkan ke dalam sel hewan. Karbohidrat yang dihasilkan oleh proses fotosintesis bahkan didistribusikan sebagai sumber nutrisi ke seluruh organisme. Meskipun kloroplas tidak tumbuh dan membelah, organel-organel tersebut terus berfungsi dalam sitoplasma sel hewan selama beberapa bulan, dan organisme inang ini disebut sebagai “hewan hijau”.
Dalam analisis terakhir, gagasan kami tentang bagaimana sel eukariotik memperoleh susunan organel kompleks mereka selama miliaran tahun evolusi harus tetap spekulatif karena peristiwa yang sedang dipertimbangkan tidak dapat diakses oleh eksperimen laboratorium langsung. Namun, satu kekuatan dari peran endosimbiosis yang diusulkan dalam asal usul evolusi organel eukariotik adalah bahwa ia melibatkan interaksi dan peristiwa yang diamati dalam sel kontemporer.