Komunikasi antar Sel
Sel saling berkomunikasi dan berinteraksi untuk menjalankan proses kehidupan. Dalam komunikasi tersebut, komitmen antar sel menjadi hal sangat penting karena keberadaan suatu sel dalam jaringan diperlukan oleh sel yang lainnya. Apabila keberadaan suatu sel sudah tidak dibutuhkan lagi dalam jaringan, misalkan dalam jaringan suatu kebutuhan sel sudah mencukupi, maka sel tersebut akan mengalami apoptosis. Hal tersebut merupakan kondisi yang perlu dilakukan pada bagian rangka tubuh dalam menjalankan fungsi dari bagian jaringan atau organ untuk mendukung proses kehidupan organisme. Jaringan komunikasi antara satu sel dengan sel yang lainnya akan menghasilkan suatu koordinasi agar dapat mengatur pertumbuhan, osmoregulasi, reproduksi dan hal lainnya pada berbagai jaringan maupun organ.
Prinsip Umum Komunikasi Sel
Komunikasi sel merupakan suatu mekanisme dimana satu sel dapat mempengaruhi sifat sel lainnya yang juga terjadi pada organisme uniseluler sebelum munculnya organisme multiseluler. Hal tersebut dibuktikan oleh penelitian yang dilakukan terhadap yeast sebagai organisme eukatiotik. Sel-sel pada yeast dalam menjalankan aktivitas hidupnya dapat berjalan secara mandiri, mereka dapat berkomunikasi dan bekerjasama dalam persiapan untuk perkawinan seksual. Saccharomyces cerevisiae misalnya, ketika sel haploid telah siap untuk kawin, maka ia akan mengeluarkan peptida (sebagai faktor kawin) yang akan memberikan sinyal pada sel-sel lawan jenis untuk berhenti berkembang biak dan mempersiapkan diri untuk kawin (Gambar 1). Fusi selanjutnya dari dua sel haploid jenis kawin yang berlawanan akan menghasilkan sel diploid, yang kemudian dapat menjalani proses meiosis dan bersporulasi, menghasilkan sel-sel haploid dengan bermacam-macam gen baru.
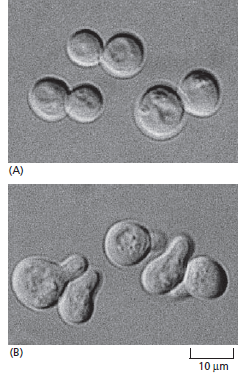
Gambar 1. Sel yeast yang merespons faktor kawin
Studi tentang yeast mutan yang tidak dapat melakukan perkawinan telah mengidentifikasi banyak protein yang diperlukan dalam proses sinyal. Protein ini membentuk jaringan pensinyalan yang mencakup protein reseptor pada permukaan sel seperti protein pengikat GTP dan protein kinase, yang masing-masing memiliki kerabat dekat pada protein yang melakukan pensinyalan terhadap sel hewan. Namun, melalui duplikasi dan divergensi gen, sistem pensinyalan pada hewan jauh lebih rumit dibandingkan pada yeast.
Molekul Sinyal Ekstraseluler Pengikat Reseptor Spesifik
Dalam melakukan komunikasi antara satu sel dengan yang lainnya untuk proses kawin, sel yeast mengeluarkan beberapa jenis peptida kecil. Sedangkan pada sel-sel hewan tingkat tinggi dalam melakukan komunikasi melalui ratusan jenis molekul sinyal, seperti peptida kecil, protein, asam amino, nukleotida, steroid, retinoid, turunan asam lemak dan bahkan gas terlarut seperti oksida nitrat dan karbon monoksida. Sebagian besar molekul sinyal ini disekresikan melalui eksositosis dari sel pemberi sinyal ke bagian ekstraseluler, sedangkan yang lainnya dilepaskan melalui proses difusi pada membran plasma. Reseptor merupakan protein spesifik yang secara khusus dapat mengikat molekul sinyal kemudian dapat merespons sel target. Molekul sinyal ekstraseluler bekerja pada konsentrasi yang sangat rendah (biasanya ≤ 10-8 M) dan reseptor yang mengenalinya biasanya mengikat mereka dengan afinitas tinggi (konstanta afinitas Ka ≥ 108 liters/mol). Dalam kebanyakan kasus, reseptor tersebut merupakan protein transmembran pada permukaan sel target. Ketika reseptor tersebut mengikat molekul sinyal ekstraseluler (ligan), reseptor menjadi aktif dan menghasilkan kaskade sinyal intraseluler yang mengubah sifat sel. Dalam kasus lain, reseptor yang berada pada sel target dan molekul sinyal harus memasuki sel untuk mengaktifkannya. Hal tersebut mengharuskan molekul sinyal yang berukuran kecil dan bersifat hidrofobik agar berdifusi melintasi membran plasma (Gambar 2).

Gambar 2. Pengikatan molekul sinyal ke reseptor permukaan sel atau reseptor intraseluler
Sebagian besar molekul sinyal bersifat hidrofilik sehingga tidak dapat melintasi membran plasma secara langsung. Sebaliknya, mereka dapat mengikat reseptor permukaan sel, yang pada gilirannya menghasilkan satu atau lebih sinyal di dalam sel target. Molekul sinyal kecil dapat berdifusi melintasi membran plasma dan berikatan dengan reseptor di dalam sel target, baik di sitosol maupun di nukleus. Banyak dari molekul sinyal kecil ini bersifat hidrofobik dan hampir tidak larut dalam air, oleh karena itu mereka diangkut dalam aliran darah dan cairan ekstraseluler lainnya setelah berikatan dengan protein pembawa (Gambar 2).
Molekul Sinyal Ekstraseluler (Bertindak pada Jarak Pendek atau Jauh)
Banyak molekul sinyal tetap terikat pada permukaan sel pensinyalan dan hanya dapat mempengaruhi sel-sel yang menghubunginya saja (Gambar 15-4A). Pensinyalan seperti itu sangat penting selama perkembangan dan dalam respons imun. Molekul yang disekresikan dapat dibawa pada jarak yang jauh untuk bekerja pada target yang jauh atau mereka dapat bertindak sebagai mediator lokal yang dapat mempengaruhi sel-sel di lingkungan secara langsung dari sel pensinyalan. Proses terakhir ini disebut pensinyalan parakrin (Gambar 15-4B). Agar sinyal parakrin dikirim hanya kepada sel target yang tepat, molekul yang disekresikan tidak boleh dibiarkan berdifusi terlalu jauh, sehingga molekul cepat diambilalih oleh sel target tetangga, dihancurkan oleh enzim ekstraseluler atau diimobilisasi oleh matriks ekstraseluler.
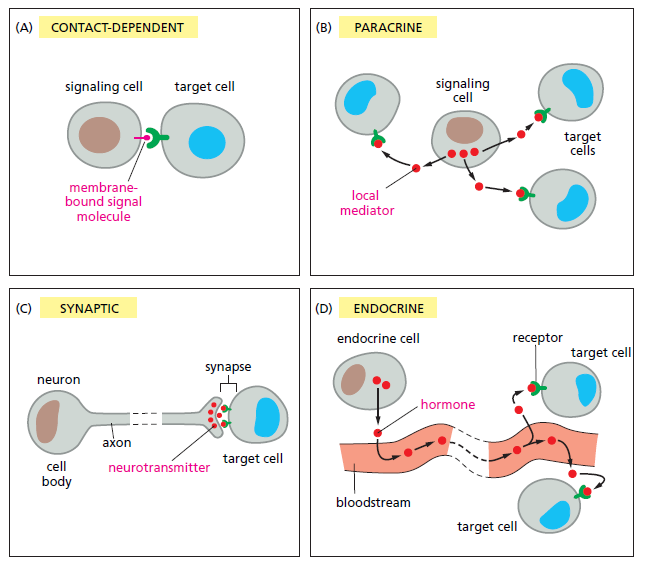
(A) Pensinyalan yang bergantung pada kontak (membutuhkan sel untuk berada dalam kontak langsung membran-membran). (B) Pensinyalan parakrin bergantung pada sinyal yang dieksresikan ke bagian ekstraseluler dan bekerja secara lokal pada sel tetangga. (C) Pensinyalan sinaptik dilakukan oleh neuron yang mentransmisikan sinyal secara elektrik di sepanjang aksonnya dan melepaskan neurotransmiter pada sinapsis (D) Pensinyalan endokrin bergantung pada sel-sel endokrin yang mensekresi hormon ke dalam aliran darah, kemudian didistribusikan secara luas ke seluruh tubuh.
Gap Junctions Memungkinkan Informasi Pensinyalan yang ditransfer oleh Sel Tetangga
Melalui gap junction aktivitas dari sel tetangga dapat terkoordinasikan. Gap juntion merupakan sambungan sel-sel khusus yang dapat terbentuk di antara membran plasma yang berdekatan dan secara langsung dapat menghubungkan sitoplasma dari sel-sel yang bergabung melalui saluran sempit. Saluran tersebut memungkinkan terjadinya pertukaran molekul sinyal intraseluler yang kecil (mediator intraseluler), seperti Ca2+ dan AMP siklik, tetapi tidak untuk makromolekul, seperti protein atau asam nukleat. Dengan demikian, sel-sel yang dihubungkan oleh gap junction dapat berkomunikasi satu sama lain secara langsung, tanpa harus melewati pembatas yang diberikan oleh membran plasma.
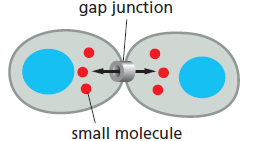
Gambar 4. Sinyal-sinyal yang melalui gap junction
Tiga Kelas Terbesar Protein Reseptor pada Permukaan Sel
Kelas terbesar protein reseptor pada permukaan sel ialah Ion-Channel-linked, G-Protein-linked dan Enzyme-linked Reseptor. Semua molekul sinyal yang larut dalam air (termasuk neurotransmiter dan semua protein sinyal) mengikat protein reseptor spesifik pada permukaan sel target yang mereka pengaruhi. Protein reseptor permukaan sel bertindak sebagai transduser sinyal. Potein reseptor mengubah proses pengikatan ligan ekstraseluler menjadi sinyal intraseluler yang mengubah sifat sel target. Sebagian besar protein reseptor permukaan sel termasuk dalam salah satu dari tiga kelas besar yang ditentukan oleh mekanisme transduksi yang digunakan protein tersebut.
Reseptor terkait saluran ion dikenal sebagai reseptor ionotropik yang terlibat pensinyalan sinaptik secara cepat antara sel yang tereksitasi secara elektrik. Pensinyalan tersebut dimediasi oleh neurotransmiter dengan jumlah yang sedikit yang berperan secara sementara untuk membuka atau menutup saluran ion yang dibentuk oleh protein pengikatnya. Kemudian dengan cara singkat dapat mengubah permeabilitas ion membran plasma sehingga sel pascasinaps tereksibilitas.
Reseptor terkait protein-G bertindak secara tidak langsung untuk mengatur aktivitas protein target yang terikat membran plasma yang terpisah. Reseptor dan protein target saling berinteraksi satu sama yang diperantarai oleh protein pengikat GTP trimerik (protein G) (Gambar 15-15B). Jika protein target adalah enzim, maka aktivitas protein tersebut dapat mengubah konsentrasi mediatror intraseluler. Akan tetapi, jika protein target merupakan saluran ion, maka aktivitas protein tersebut dapat mengubah permeabilitas ion membran plasma.
Reseptor terkait-enzim, ketika diaktifkan berperan secara langsung sebagai enzim. Reseptor tersebut dibentuk oleh protein transmembran single-pass yang memiliki situs pengikatan ligan di luar sel dan pengikatan enzim di dalam. Reseptor terkait-enzim memiliki struktur yang heterogen dibandingkan dengan dua kelas protein reseptor lainnya.
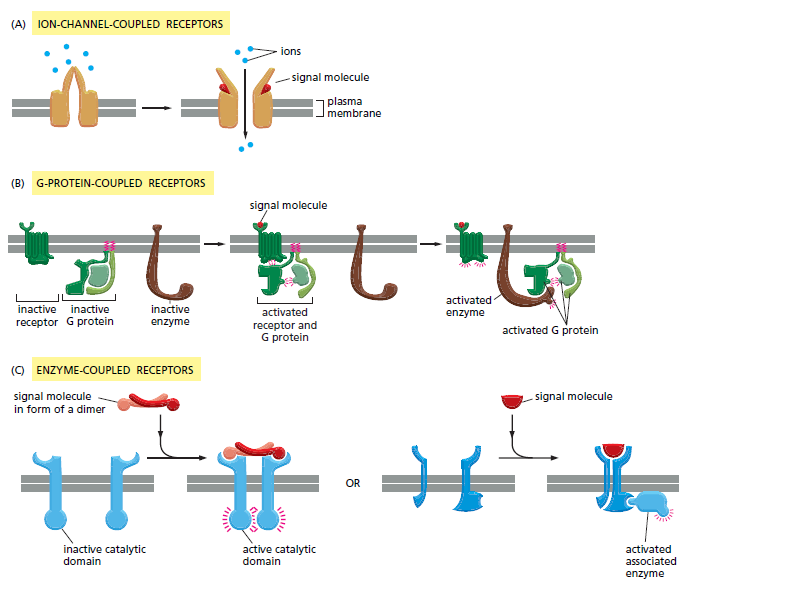
Gambar 5. Tiga kelas reseptor permukaan sel: (A) Reseptor terkait saluran ion, (B) reseptor terkait protein G dan (C) reseptor terkait enzim. Meskipun banyak reseptor terkait-enzim memiliki aktivitas enzim intrinsik, seperti yang ditunjukkan di sebelah kiri, banyak lainnya bergantung pada enzim terkait, seperti yang ditunjukkan di sebelah kanan.
Reseptor Permukaan Sel yang sering Diaktifkan
Reseptor permukaan sel yang sering diaktifkan akan menyampaikan sinyalnya melalui molekul kecil dan jaringan protein pensinyalan intraseluler. Sinyal yang diterima permukaan sel baik oleh reseptor terkait protein-G ataupun reseptor terkait-enzim kemudian diteruskan ke bagian dalam sel melalui kombinasi molekul pemberi sinyal intraseluler besar dan kecil. Rantai yang dihasilkan dari proses pensinyalan intraseluler kemudian mengubah protein target, yang bertanggung jawab dalam modifikasi sifat sel. Molekul pensinyalan intraseluler kecil disebut sebagai perantara intraseluler kecil atau pembawa pesan kedua, sedangkan pembawa pesan pertama merupakan sinyal ekstraseluler. Sinyal intraseluler dan ekstraseluler dihasilkan dalam jumlah yang besar sebagai respons terhadap aktivasi reseptor. Keduanya dapat menyebar menjauhi sumbernya dengan sangat dan memancarkan sinyal ke bagian lain dari sel. AMP siklik dan Ca2+ dapat larut dalam air dan berdifusi ke dalam sitosol, sedangkan yang lainnya, seperti diasilgliserol dapat larut dalam lemak dan berdifusi ke dalam membran plasma. Sebagian besar molekul pensinyalan intraseluler ialah protein pensinyalan intraseluler yang membantu dalam menyampaikan sinyal ke dalam sel dengan menghasilkan perantara intraseluler kecil atau mengaktifkan protein pensinyalan. Protein tersebut dapat diklasifikasikan menurut fungsi khususnya (Gambar 6).
- Relay protein –> hanya akan meneruskan pesan ke komponen sinyal berikutnya yang berada dalam rantai.
- Protein pembawa pesan –> membawa sinyal dari satu bagian ke bagian yang lain dari sel, seperti dari sitosol ke nukleus.
- Protein adaptor –> menghubungkan satu protein pensinyalan ke protein pensinyalan lainnya.
- Protein penguat –> yaitu dapat berupa enzim atau saluran ion. Protein tersebut dapat meningkatkan sinyal yang mereka terima, baik dengan memproduksi perantara intraseluler kecil dalam jumlah besar atau dengan cara mengaktifkan protein sinyal intraseluler hilir dalam jumlah yang besar juga.
- Protein transduser –> dapat mengubah sinyal menjadi bentuk yang berbeda. Contohnya ialah enzim yang membuat AMP siklik: ia dapat mengubah sinyal dan menguatkannya, sehingga dapat berperan sebagai transduser dan penguat.
- Protein bifurkasi –> menyebarkan sinyal dari satu jalur pensinyalan ke jalur lainnya.
- Protein integrator –> dapat menerima sinyal dari dua atau lebih jalur pensinyalan dan mengintegrasikannya sebelum meneruskan sinyal.
- Protein pengatur gen laten –> diaktifkan pada reseptor permukaan sel kemudian akan bermigrasi ke nukleus untuk merangsang transkripsi gen.
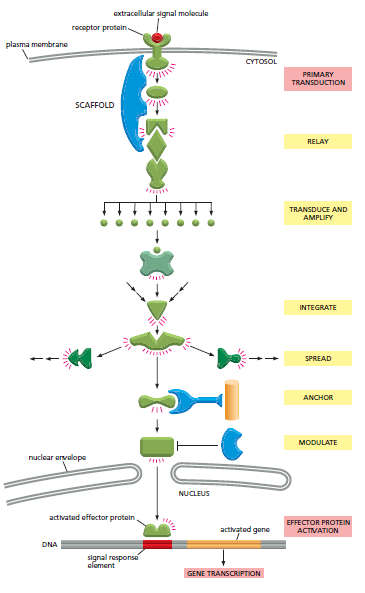
Gambar 6. Berbagai jenis protein pensinyalan intraseluler di sepanjang jalur pensinyalan dari reseptor permukaan sel ke nukleus
Gambar di atas merupakan serangkaian protein pensinyalan dan intraseluler kecil sebagai perantara yang menyampaikan sinyal ekstraseluler ke dalam sel. Hal tersebut dapat menyebabkan perubahan ekspresi gen. Sinyal yang telah diperkuat dapat ditransduksikan (diubah) kemudian didistribusikan selama dalam perjalanan. Sebagian besar langkah (tahap) dapat dimodulasi oleh sinyal ekstraseluler dan intraseluler lainnya, sehingga hasil akhir dari satu sinyal sangat bergantung pada faktor lain yang mempengaruhi sel. Sehingga pada akhirnya, jalur pensinyalan dapat mengaktifkan atau menonaktifkan protein target yang dapat mengubah sifat sel. Pada contoh dalam Gambar 6 yang menjadi target ialah protein pengatur gen.
Pensinyalan Autokrin
Pensinyalan autokrin dapat mengkoordinasikan suatu keputusan berdasarkan kelompok sel identik. Sebagian bentuk pensinyalan yang dibahas sebelumnya, memungkinkan satu sel dapat mempengaruhi sel yang lainnya, dimana sel pensinyalan dan target merupakan tipe sel yang berbeda. Padahal, suatu sel dapat mengirimkan sinyal ke sel yang lain dari jenis sel yang sama serta untuk sel itu sendiri. Pensinyalan autokrin yang seperti itu, sel mengeluarkan molekul sinyal yang dapat mengikat kembali reseptornya sendiri. Selama proses perkembangan, sel yang telah diinstruksikan ke jalur differensiasi tertentu, kemungkinan telah mengeluarkan sinyal autokrin untuk diri sel tersebut. Pensinyalan autokrin paling efektif bila dilakukan secara simultan oleh sel-sel tetangga dari jenis yang sama dan nantinya akan digunakan untuk mendukung kelompok sel yang identik (sejenis) agar membuat keputusan perkembangan yang sama. Dengan demikian, pensinyalan autokrin dianggap sebagai salah satu mekanisme yang mungkin mendasari “efek komunitas” yang mulai diamati pada perkembangan awalnya, di mana sekelompok sel identik dapat merespons sinyal pemicu diferensiasi tetapi satu sel yang terisolasi dari jenis yang sama tidak dapat merenspons sinyal tersebut.
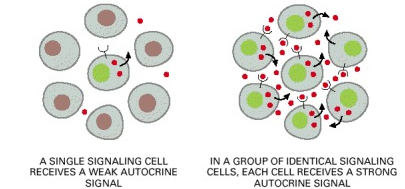
Gambar 7. Pensinyalan autokrin. Sekelompok sel yang identik akan menghasilkan konsentrasi sinyal yang disekresikannya lebih tinggi daripada konsentrasi sel tunggal. Ketika sinyal ini mengikat kembali ke reseptor pada jenis sel yang sama, akan mendorong sel tersebut untuk merespons sebagai sebuah kelompok yang terkoordinir.
Pemrograman sel untuk menanggapi Kombinasi Spesifik dari Molekul Sinyal Ekstraseluler
Sel yang khusus pada organisme multiseluler akan terpaut pada ratusan sinyal yang berbeda di lingkungannya. Sinyal-sinyal tersebut dapat larut, terikat pada matriks ekstraseluler atau terikat pada permukaan sel tetangga dan nantinya akan berkombinasi satu dengan yang lainnya. Sel harus menanggapi respons sinyal secara selektif, sesuai dengan karakter spesifiknya sendiri, yang diperolehnya melalui spesialisasi sel progresif dalam proses perkembangannya. Sebuah sel dapat diprogram untuk menanggapi satu kombinasi sinyal dengan membedakan kombinasi lain, dengan mengalikan dan dengan melakukan beberapa fungsi khusus seperti kontraksi atau sekresi. Sebagian besar sel yang terdapat pada hewan tingkat tinggi juga diprogram untuk bergantung pada kombinasi sinyal tertentu untuk bertahan hidup. Ketika kehilangan sinyal tersebut (dalam cawan kultur, misalnya), sel akan mengaktifkan program bunuh diri dan membunuh dirinya sendiri yang disebut sebagai kematian sel yang terprogram, atau apoptosis (Gambar 8). Karena jenis sel yang berbeda akan memerlukan kombinasi sinyal untuk kelangsungan hidup, dimana setiap jenis sel dibatasi pada lingkungan yang berbeda di dalam tubuh. Kemampuan untuk menjalani apoptosis merupakan sifat dasar sel hewan.
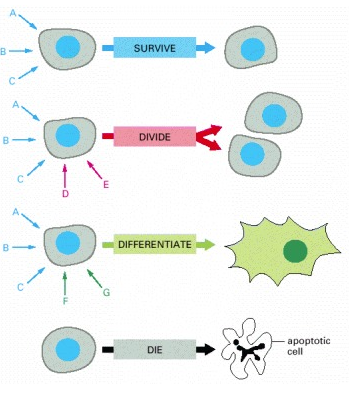
Gambar 8. Setiap Sel Diprogram untuk Menanggapi Kombinasi Spesifik dari Molekul Sinyal Ekstraseluler. Setiap jenis sel akan menampilkan satu set reseptor yang nantinya untuk merespons serangkaian molekul sinyal yang dihasilkan oleh sel lain. Molekul sinyal tersebut saling kombinasi untuk mengatur sifat dari sel. Sel individu membutuhkan banyak sinyal untuk bertahan (panah biru) dan sinyal tambahan untuk membelah (panah merah) atau membedakan (panah hijau). Jika kehilangan sinyal untuk kelangsungan hidup, maka sel akan mengalami kematian sel terprogram atau apoptosis.
Jenis sel yang berbeda dapat merespon secara serbeda pula terhadap molekul sinyal ekstraseluler yang sama. Secara spesifik, sel akan bereaksi secara beragam terhadap lingkungannya. Keragaman reaksi tersebut sesuai dengan set protein reseptor yang dimiliki oleh sel. Set protein tersebutlah yang menentukan subset tertentu dari sinyal yang dapat ditanggapinya dan bervariasi sesuai dengan mesin intraseluler. Dimana sel tersebut akan mengintegrasikan dan menterjemahkan sinyal yang diterimanya (lihat Gambar 9 ). Dengan demikian, molekul sinyal tunggal akan memiliki dampak yang berbeda pada sel target yang berbeda pula. Neurotransmitter asetilkolin, misalnya, akan merangsang kontraksi sel otot rangka, tetapi juga menurunkan kecepatan dan kekuatan kontraksi sel otot jantung. Hal tersebut dikarenakan protein reseptor asetilkolin pada sel otot rangka berbeda dengan yang ada pada sel otot jantung. Tetapi perbedaan reseptor tersebut tidak selalu menjelaskan efek (dampak) yang berbeda. Dalam banyak kasus, molekul sinyal yang sama akan berikatan dengan protein reseptor yang sama (identik), namun juga menghasilkan respons yang berbeda dalam berbagai jenis sel target. Hal tersebut mencerminkan perbedaan dalam mesin internal tempat reseptor digabungkan (Gambar 10).
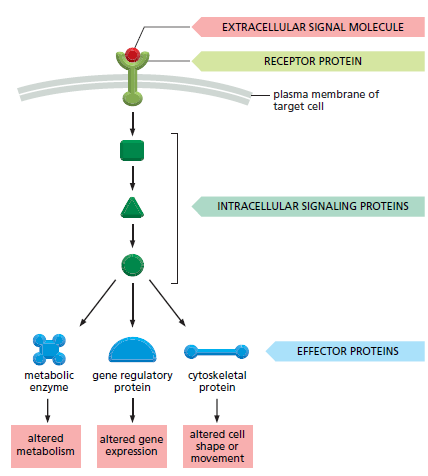
Gambar 9. Jalur pensinyalan intraseluler sederhana yang diaktifkan oleh molekul sinyal ekstraseluler. Biasanya, molekul sinyal akan mengikat protein reseptor yang tertanam dalam membran plasma sel target serta mengaktifkan satu atau lebih jalur pensinyalan intraseluler yang diperantarai oleh serangkaian protein pensinyalan. Akhirnya, satu atau lebih protein pensinyalan intraseluler mengubah aktivitas protein efektor dan sifat sel tersebut.
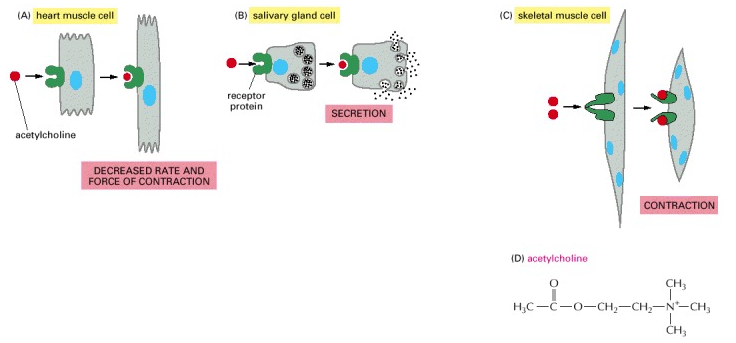
Gambar 10. Berbagai respons yang diinduksi oleh neurotransmitter asetilkolin. Jenis sel yang berbeda terspesialisasi untuk merespon asetilkolin dengan cara yang berbeda pula. (A dan B) asetilkolin berikatan dengan protein reseptor yang serupa, tetapi sinyal intraseluler yang dihasilkan diinterpretasikan secara berbeda dalam sel yang terspesialisasi untuk fungsi yang berbeda juga. (C) sel otot menghasilkan jenis protein reseptor yang berbeda untuk asetilkolin kemudian menghasilkan sinyal intraseluler yang berbeda dari reseptor yang ditunjukkan pada (A) dan (B) serta menghasilkan efek yang berbeda. (D) Struktur kimia asetilkolin.

Informasi ini menarik
Good providers