Fotosintesis
Fotosintesis merupakan sebuah proses seluler tumbuhan yang sangat penting, baik bagi tumbuhan itu sendiri, maupun makhluk lain di muka bumi (termasuk manusia). Secara sederhana, fotosintesis dapat dijelaskan sebagai suatu proses pengambilan karbon dioksida (CO2) dan air (H2O) dari lingkungan, yang mana nantinya akan menghasilkan oksigen (O2) dan pati (C6H12O6), dengan bantuan cahaya matahari. Sementara itu, literature lainnya menyebutkan bahwa fotosintesis dapat diartikan sebagai konversi energi cahaya menjadi energi kimia yang disimpan dalam bentuk glukosa dan molekul organik lainnya. Lantas, manakah pendapat yang benar mengenai fotosintesis? Agar lebih paham, mari ikuti penjelas berikut mengenai kloroplas, cahaya dan proses fotosintesis beserta evolusinya.
Kloroplas: Pusat Fotosintesis di Tumbuhan
Kloroplas merupakan organel yang khusus terdapat pada organisme autotrof (sel tumbuhan dan beberapa jenis alga) yang berperan penting dalam proses fotosintesis. Ditinjau dari sisi evolusi, teori endosimbion menyatakan bahwa kloroplas awalnya merupakan organisme prokariotik fotosintetik (misalnya cyanobacteria) yang bersimbiosis di dalam nenek moyang sel organisme eukariotik Bukti kuat yang mendukung teori ini adalah adanya materi genetik tersendiri yang dimiliki oleh kloroplas, yang berbeda dengan materi genetik di inti sel.
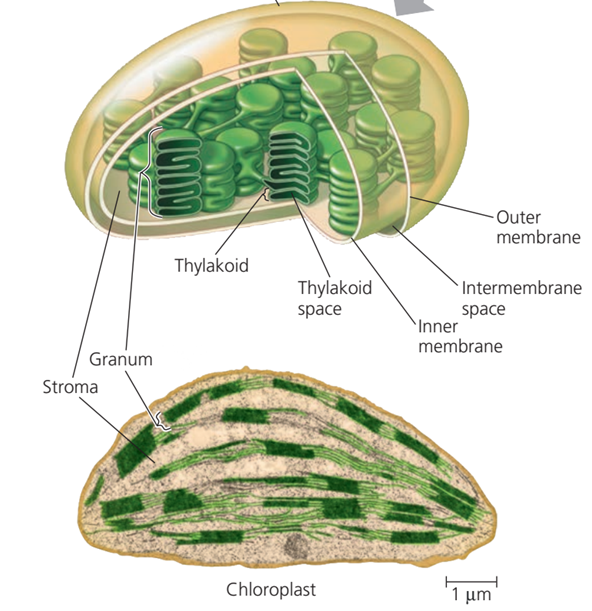
Gambar. Struktur Kloroplas
Seluruh bagian tumbuhan yang berwarna hijau, baik itu batang yang masih muda ataupun buah yang belum matang mengandung kloroplas. Namun, kloroplas paling banyak terdapat di jaringan mesofil daun, yaitu lapisan jaringan yang berada tepat di bawah epidermis daun. Kloroplas merupakan salah satu contoh organel sel dengan membran ganda (double-membrane-bounded organelle), terdiri dari membran luar (outer membrane) dan membran dalam (inner membrane). Kedua membran dipisahkan oleh ruang antar-membran (intermembrane space) yang sangat berperan saat proses fotosintesis. Membran dalam dipenuhi oleh substansi cair transparan yang disebut dengan stroma. Di dalam stroma, terkandung sebuah sistem bermembran lagi yang di sebut tilakoid. Pada beberapa spesies tumbuhan, tilakoid akan tersusun bertumpuk dan tumpukan ini di sebut sebagai granum (jamak: grana).
Cahaya yang Efektif untuk Fotosintesis
Apabila dikaji dari segi bahasa, fotosintesis atau photosynthesis berasal dari dua kata yaitu, photo (cahaya) dan synthesis (pembentukan). Secara sederhana, proses ini dapat kita artikan sebagai sebuah reaksi pembentukan yang berbantukan cahaya, dalam hal ini adalah cahaya matahari. Dari sini, dapat dipahami bahwa cahaya merupakan elemen penting dalam fotosintesis. Secara fisika, cahaya matahari merupakan energi dalam bentuk gelombang elektromagnetik. Gelombang elektromagnetik ini, kemudian dapat diklasifikasikan berdasarkan beberapa bagian lagi berdasarkan panjang gelombangnya (spectrum elektromagnetik). Cahaya tampak, yaitu gelombang elektromagnetik yang dapat ditangkap mata manusia, memiliki panjang gelombang antara 38—750 nm. Semakin besar nilai panjang gelombangnya, amka akan semakin kecil energinya, begitupun sebaliknya.
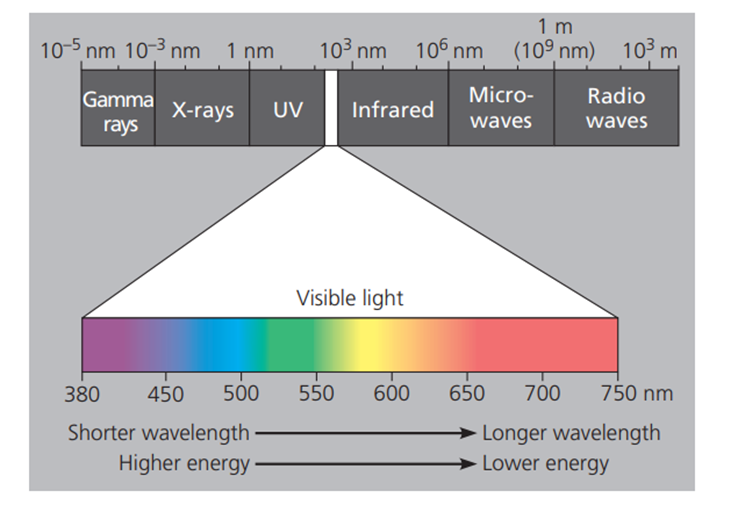
Gambar. Spektrum elektromagnetik dari cahaya tampak
Yang menjadi pertanyaan sekarang adalah, apakah proses fotosintesis dapat memanfaatkan seluruh jenis cahaya tampak? Atau hanya akan berjalan efektif jika menggunakan cahaya dengan panjang gelombang tertentu saja? Untuk menjawab pertanyaan ini, kita dapat merujuk pada percobaan yang dilakukan Theodor Engelmann pada tahun 1883. Engelmann, menggunakan ganggang Spyrogyra sp. yang berbentuk filamen yang disinari seberkas cahaya tampak yang dilewatkan melalui prisma. Dengan adanya cahaya, maka Spyrogyra akan melangsungkan fotosintesis. Engelmann kemudian menggunakan bakteri aerobik untuk mendeteksi pada panjang gelombang berapa fotosintesis paling efektif berjalan. Bakteri aerobik akan berkumpul pada daerah degan konsentrasi oksigen tinggi, yang mana oksigen tinggi menandakan bahwa laju fotosintesis di daerah itu juga tinggi. Hasil percobaan menunjukkan bahwa bakteri aerobik paling banyak berkumpul pada daerah ganggang yang disinari cahaya ungu-biru dan merah.
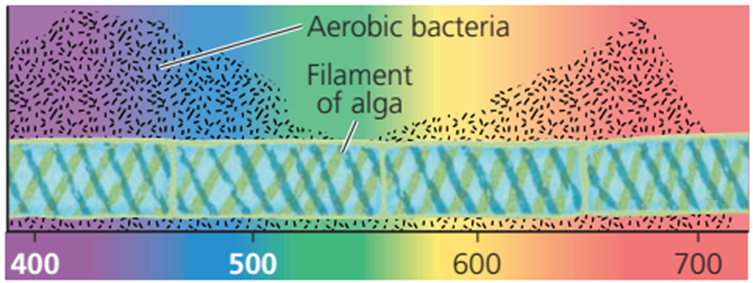
Gambar. Percobaan Engelman
Saat membicarakan cahaya sebagai gelombang, banyak sifat dari cahaya itu sendiri yang dapat dijelaskan, namun dalam kondisi tertentu cahaya dapat dijelaskan sebagai sesuatu yang seolah-olah terdiri dari partikel diskrit. Partikel diskrit ini kemudian disebut sebagai foton (photon). Foton bukanlah objek nyata, tetapi foton bertindak selayaknya objek yang masing-masing memiliki jumlah energi yang tetap. Sebagaimana yang teLah dijelaskan sebelumnya, jumlah energi berbanding terbalik dengan panjang gelombang cahaya: semakin pendek panjang gelombang, semakin besar energi setiap foton cahaya itu. Jadi, foton cahaya ungu mengemas energi hampir dua kali lebih banyak daripada foton cahaya merah.
Ketika cahaya bertemu dengan benda lain, cahaya dapat dipantulkan, ditransmisikan ataupun diserap. Substansi yang mampu menyerap cahaya disebut sebagai pigmen. Pigmen yang berbeda akan menyerap cahaya dengan panjang gelombang yang berbeda pula. Bila suatu pigmen menyerap cahaya dengan gelombang tertentu, warna tersebut akan menghilang dan warna yang tampak adalah warna yang dipantulkan. Kloroplas sebagai pigmen fotosintetik akan menyerap cahaya ungu-biru dan merah. Melalui gambar 3, tampak bahwa cahaya hijau bukanlah cahaya yang efektif untuk fotosintesis, sehingga cahaya ini dipantulkan. Itulah mengapa daun secara umum terlihat berwarna hijau.
Proses Fotosintesis
Proses fotosintesis secara umum dapat diklasifikasikan dalam dua tahapan, yakni reaksi terang (light-dependent reaction) dan reaksi gelap (light independent reaction). Seperti namanya, keberlangsungan reaksi terang bergantung pada keberadaan cahaya, namun tidak untuk reaksi gelap. Reaksi gelap dapat terjadi pada kondisi ada atau tidak ada cahaya, asalkan komponen yang dibutuhkan untuk memulai reaksi ini (produk hasil reaksi terang) tersedia.
Reaksi Terang
Reaksi terang fotosintesis dimulai ketika radiasi cahaya matahari mulai menyentuh daun dan energi foton mulai diserap oleh klorofil. Penyerapan energi foton di klorofil mengakibatkan elektron yang terdapat pada kloroplas berpindah (bereksitasi) ke daerah dengan energi potensial yang lebih tinggi. Setelah tereksitasi, elektron sebagaimana molekul berenergi tinggi lain bersifat sangat tidak stabil. Oleh karenanya, dalam waktu sepersekian detik, elektron akan kembali ke posisi semula sambil melepaskan energi panas. Untuk memahami proses eksitasi elektron perhatikan gambar 4 berikut.
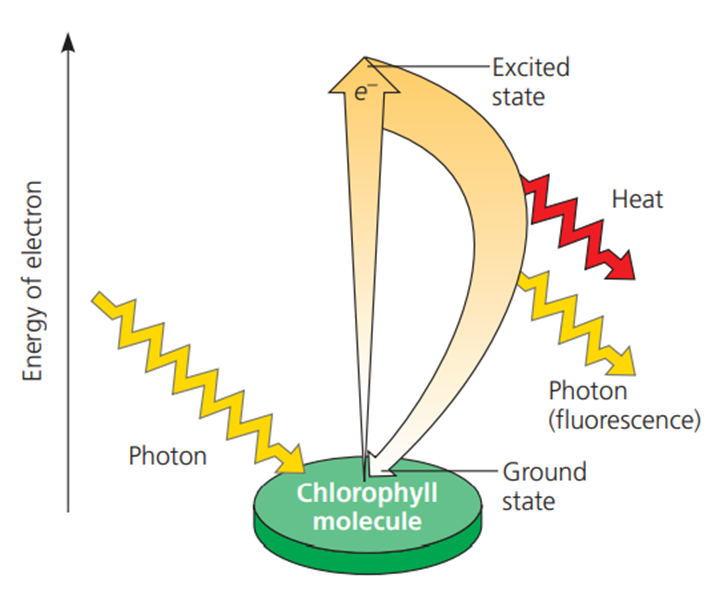
Gambar. Eksitasi elektron oleh energi cahaya
Pigmen klorofil yang berada di membran tilakoid, terorganisasi dengan molekul dan protein lainnya membentuk sebuah struktur yang disebut sabagai fotosistem. Sebuah fotosistem terdiri dari kompleks pusat reaksi (reaction-center complex) yang dikelilingi oleh beberapa kompleks pemanen cahaya (light-harvesting complex). Kompleks pusat reaksi adalah kumpulan protein yang terorganisir yang memuat beberapa molekul seperti pasangan molekul klorofil a dan akseptor elektron primer (primary elektron acceptor). Sementara itu, kompleks pemanen cahaya terdiri dari berbagai molekul pigmen seperti klorofil a, klorofil b, dan karotenoid. Dengan adanya variasi jenis dan jumlah pigmen ini, fotosistem dapat mengumpulkan cahaya dengan panjang gelombang dan permukaan yang lebih luas lagi. Kompleks pemanen cahaya ini bertindak sebagai antena untuk kompleks pusat reaksi.
Ketika molekul pigmen menyerap foton, energi ditransfer dari molekul pigmen ke molekul pigmen dalam kompleks pemanen cahaya, hingga diteruskan ke pasangan molekul klorofil a di kompleks pusat reaksi. Pasangan molekul klorofil a dalam kompleks pusat reaksi cukup special karena lingkungan molekulnya, lokasinya, dan karena molekul-molekul lain yang saling berhubungan. Kondisi ini memungkinkan mereka untuk menggunakan energi dari cahaya tidak hanya untuk mengeksitasi elektronnya ke tingkat energi yang lebih tinggi, tetapi juga untuk mentransfernya ke molekul yang berbeda, dalam pemahasan ini ialah akseptor elektron primer.
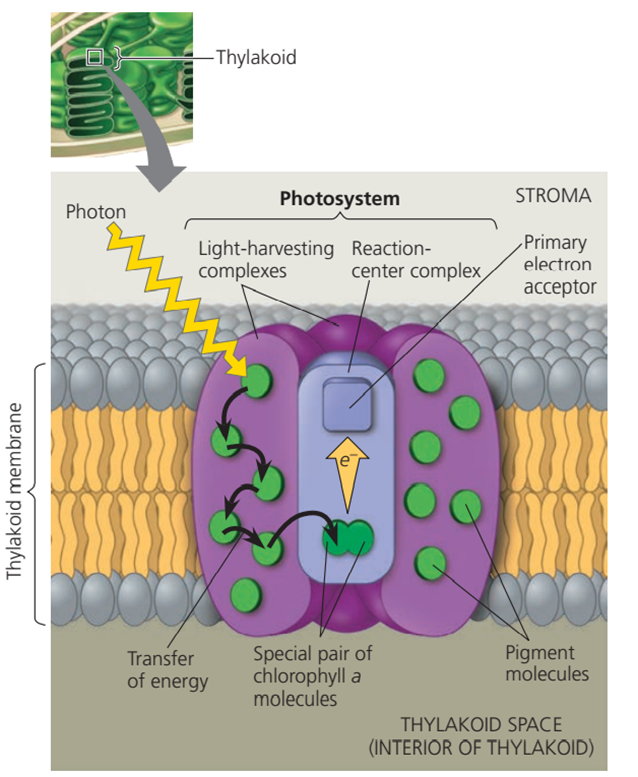
Gambar. Proses penyerapan energi cahaya di fotosistem
Terdapat dua jenis fotosistem yang berada di membran tilakoid, yaitu Fotosistem I (PS I) dan Fotosistem II (PS II). Pusat reaksi dari PS II dikenal sebagai P680 karena pasangan klorofil a yang terdapat pada fotosistem ini efektif menyerap cahaya dengan panjang gelombang 680 nm. Sementara itu, pusat reaksi PS I dikenal sebagai PS700 karena pasangan klorofil a nya efektif menyerap cahaya dengan panjang gelombang 700 nm. Penamaan fotosistem dilakukan berdasarkan mana fotosistem yang ditemukan lebih dulu, tetapi PS II pada reaksi terang akar berperan terlebih dahulu. Kedua pigmen ini, P680 dan P700, adalah molekul klorofil a yang hampir identik. Namun, hubungan mereka dengan protein yang berbeda dalam membran tilakoid mempengaruhi distribusi elektron dalam dua pigmen dan menjelaskan sedikit perbedaan dalam sifat menyerap cahaya. Reaksi terang dapat berlangsung dalam dua skenario, yaitu secara non-siklik/linear dan siklik.
Reaksi non-siklik/linear
Energi foton dari cahaya diserap masuk ke dalam Fotosistem II, tepatnya pada molekul pigmen klorofil. Seperti yang sudah dijelaskan sebelumnya, elektron akan tereksitasi, berpindah ke daerah dengan energi potensial yang lebih tinggi, lalu kembali ke kondisi awal. Setelah kembali ke kondisi awal, energi akan ditransfer ke molekul pigmen yang berdekatan dan memicu terjadinya eksitasi elektron. Panah berwarna hita, pada Gambar 5 menunjukkan aliran energi yang ditransfer dari satu pigmen ke pigmen lainnya. Proses ini terus berlangsung hingga akhirnya mencapai P680.
Elektron yang dieksitasi dari P680 kemudian akan ditangkap oleh akseptor elektron primer (primary elektron acseptor). P680 akan berubah menjadi P680+ karena telah kehilangan elektronnya. Pada saat yang bersamaan, suatu jenis enzim akan memcah molekul air yang diserap akar tumbuhan menjadi dua elektron, dua ion hidrogen (H+) dan sebuah atom oksigen. Elektron yang dilepaskan dari reaksi pemecahan air ini kemudian akan menggantikan elektron yang hilang di P680+. Dua ion hidrogen (H+) yang dihasilkan akan dialirkan ke ruang tilakoid. Sementara itu, atom oksigen akan bergbaung dengan atom oksigen hasil pemecahan molekul air lain dan akan membentuk oksigen (O2) yang akan dialirkan ke atmosfer.
Elektron yang telah ditangkap oleh akseptor elektron primer akan diteruskan ke Fotosistem I melalui rantai transport elektron, yang disusun atas elektron carrier plastoquinone (Pq), sebuah kompleks sitokrom, dan sebuah protein yang disebut sebagai plastocyanin (Pc). Seiring dengan aliran elektron yang melalui tiga komponen ini, pada tiap-tiap komponen berlangsung reaksi redoks yang melepaskan satu ion hidrogen (H+) ke ruang tilakoid. Pelepasan ion hidrogen (H+) ke ruang tilakoid akan memicu terbentuknya gradient proton melewati memban tilakoid. Melalui sebuah proses yang disebut sebagai kemiosmosis, gradient proton akan menghasilkan ATP. Setelah sampai di PS I, elektron akan langsung diterima oleh P700 dan kemudian terkesitasi dan ditangkan oleh akseptor elektron primer pada PS II. Setelah kehilangan elektron, P700 akan berubah menjadi P700+. Elektron yang hilang akan segera digantikan oleh elektron lain yang tereksitasi akibat diserapnya cahaya oleh molekul pigmen PS I. Elektron yang dterima oleh akseptor elektron primer di PS I sekali lagi akan melewati rantai transport elektron yang disusun atas sebuah protein, yakni Ferredoxin (Fd). Namun, rantai ini tidak akan menciptakan gradient proton sehingga tidak menghaislkan ATP. Elektron dari Fd akan diterima oleh NADP+ dengan bantuan enzim NAPD+ reduktase membentuk molekul NADPH. Pembentukan molekul NADPH memerlukan dua buah elektron.
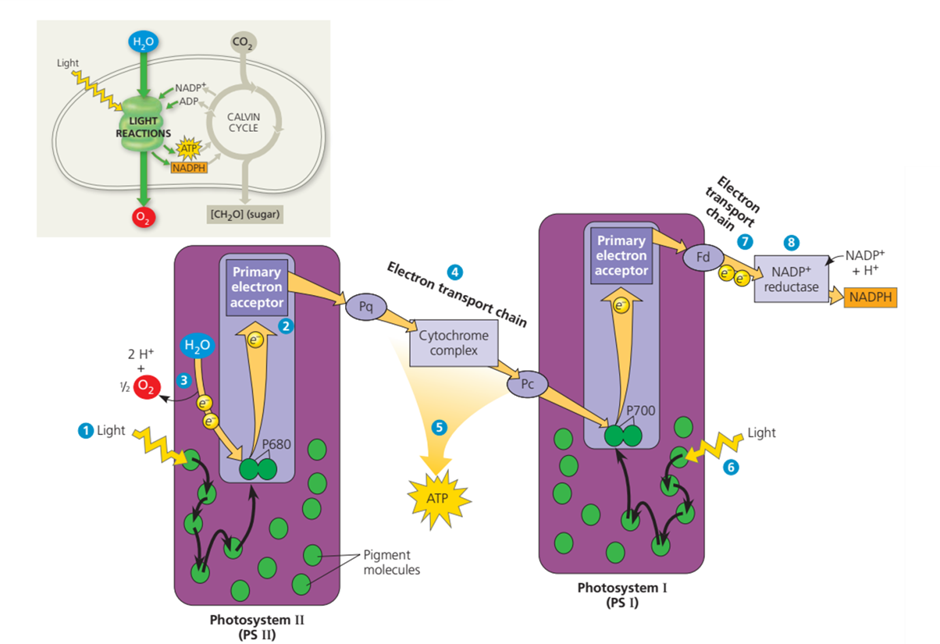
Gambar. Reaksi Non-Siklik Fotosintesis
Reaksi Siklik
Pada beberapa kondisi tertentu, ada kalanya penangkapan cahaya hanya melibatkan PS I saja. elektron yang terkesitasi dari P700 akan kembali menuju Fd, kompleks sitokrom, dan Pc kemudian berbalik menuju P700 kembali. Reaksi siklik menghasilkan ATP namun tidak menghasilkan NADPH dan tidak melepaskan oksigen ke atmosfer. Proses reaksi siklik ditampilkan pada Gambar.
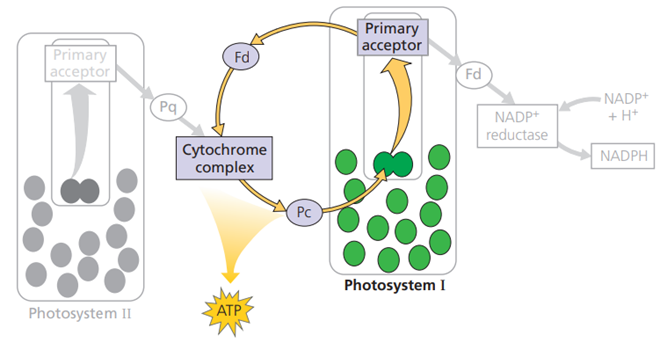
Gambar. Reaksi Siklik Fotosintesis
Beberapa kelompok bakteri fotosintetik seperti bakteri sulfur ungu dan hijau diketahui hanya memiliki satu jenis fotosistem saja. Organisme seperti ini hanya menghasilkan energi dalam bentuk ATP. Ahli evolusi menganggap bahwa organisme-organisme ini merupakan nenek moyang dari bakteri fotosintetik yang pertama berevolusi. Reaksi siklik fotosintesis juga dapat dijumpai pada organisme lain yang memiliki dua fotosistem, seperti Cyanobacteria. Meskipun dianggap sebagai haisl peninggalan proses evolusi, kemampuan untuk melangsungkan reaksi siklik dipercaya oleh para ahli mendatangkan keuntungan tertentu bagi organisme yang bersangkutan. Tanaman dengan mutasi yang membuat mereka tidak dapat melangsungkan reaksi siklik, mampu tumbuh dengan baik dalam kondisi dengan cahaya rendah, tetapi tidak tumbuh dengan baik di tempat yang terang. Ini adalah bukti bahwa reaksi siklik mungkin bersifat fotoprotektif.
Reaksi Gelap
Reaksi gelap tidak harus terjadi pada malam hari. Kunci dari keberlangsungan reaksi gelap adalah tersedianya energi hasil reaksi terang, yakni ATP dan NADPH. Inti pembahasan dari reaksi gelap adalah bagaimana karbon dioksida (CO2) yang diserap dari lingkungan dapat diubah menjadi glukosa menggunakan bantuan ATP dan NADPH. Reaksi gelap umunya disebut juga sebagai Siklus Calvin dan dapat diklasifikasikan dalam tiga tahapan, yakni tahap fiksasi karbon, tahap reduksi dan tahap regenerasi, dan terjadi di stroma.
Fiksasi Karbon
Karbon dioksida (CO2) yang ada di lingkungan, masuk ke dalam daun, tepatnya di stroma melalui stomata. Molekul CO2 akan diikat/difiksasi oleh molekul gula dengan lima atom karbon, yakni ribulose bisphosphate (RuBP). Proses pengikatan CO2 oleh RuBP dibantu oleh enzim RuBP carboxylase-oxygenase (rubisco). Pengikatan CO2 oleh RuBP akan menghasilkan molekul dengan enam atom karbon, namun sangat tidak stabil. Karenanya, molekul ini langsung terpecah menjadi dua molekul dengan tiga atom karbon, yakni 3-phosphoglycerate.
Reduksi
ATP yang dihasilkan pada reaksi terang selanjutnya akan menyumbangkan satu gugus posfat pada masing-masing 3-phosphoglycerate dan membentuk dua molekul 1,3-biphosphoglycerate. Kedua molekul 1,3-biphosphoglycerate ini kemudian kembali kehilangan satu gugus posfat sehingga membentuk dua molekul glyceraldehyde 3-phosphate (G3P). G3P merupakan gula dengan tiga atom karbon. Produk hasil fotosintesis merupakan gula dengan enam atom karbon. Walaupun terdapat dua molekul G3P (yang berarti tersedia enam atom karbon), hanya akan ada satu molekul G3P yang tereduksi untuk setiap satu atom karbon yang masuk ke dalam siklus Calvin. Satu molekul G3P lainnya harus dipakai untuk melanjutkan keberlangsungan siklus Calvin.
Degenerasi
Untuk melanjutkan siklus Calvin, sebenarnya dibutuhkan sebanyak 15 karbon (lima buah molekul G3P). Untuk itu, pada tahap awal setidaknya RuBP perlu untuk mengikat minimal tiga buah molekul CO2 agar siklus Calvin dapat tetap berlangsung. Namun, perlu diingat kembali bahwa hanya aka nada satu molekul G3P yang tereduksi untuk setiap tiga CO2 yang diikat, sehingga setidaknya harus ada enam molekul CO2 yang diikat oleh RuBP untuk menghasilkan satu molekul glukosa. Dengan demikian, ilustrasi Siklus Calvin pada Gambar 7 menunjukkan hal demikian (terdapa enam molekul CO2 yang diikat oleh RuBP). Dalam serangkaian reaksi yang kompleks, kerangka karbon dari lima molekul G3P disusun ulang pada langkah-langkah terakhir dari siklus Calvin menjadi tiga molekul RuBP. Untuk mencapai ini, dibutuhkan tiga molekul ATP. RuBP yang telah teregenerasi siap untuk menerima CO2 lagi, dan siklus berlanjut.
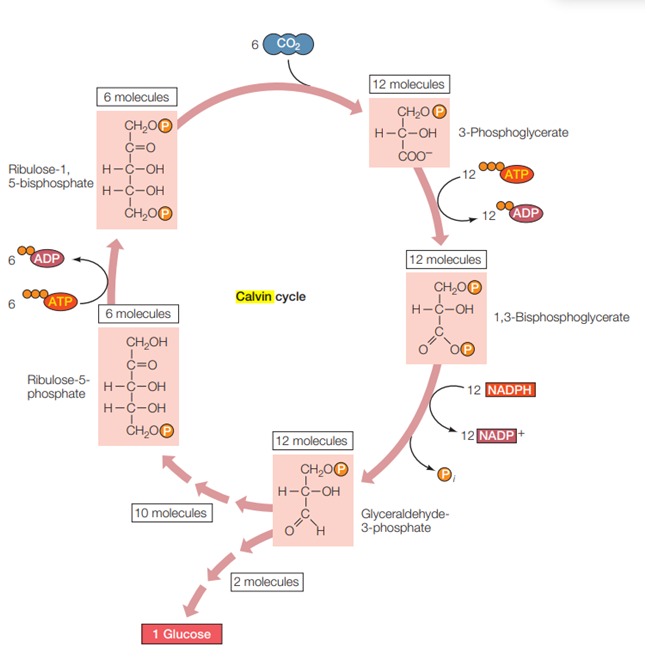
Gambar. Siklus Calvin
Tumbuhan C3, C4 dan CAM
Karbon dioksida, masuk ke dalam daun melalui pori-pori daun (stomata). Selain bertugas sebagai jalur masuk CO2, stomata juga berperan dalam proses lain, misalnya pelepasan uap air dari tumbuhan saat transpirasi berlangsung. Proses membuka dan menutupnya stomata, dipengaruhi oleh banyak hal, salah satunya adalah suhu lingkungan. Saat suhu lingkungan tinggi, stomata umumnya menutup untuk mencegah kehilangan air akibat transpirasi. Dengan menutupnya stomata, konsentrasi CO2 di dalam sel-sel daun akan berkurang dan dapat menyebabkan suatu fenomena yang disebut dengan fotorespirasi. Fotorespirasi adalah kondisi di mana RuBP yang seharusnya mengikat CO2 malah mengikat O2 akibat konsentrasi oksigen yang sangat tinggi di daun sementara konsentrasi karbon dioksida rendah. Tingginya konsentrasi O2, disebabkan oleh siklus terang yang terus menerus berlangsung yang menghasilkan oksigen, sementara Siklus Calvin tidak kunjung berlangsung karena CO2 yang terbatas. Fotorespirasi dapat dikatakan sebagai hal yang sia-sia, karena produk akhir fotosintesis, yaitu gula tidak terbentuk.
Namun, beberapa jenis tumbuhan memiliki mekanisme tertentu untuk mencegah terjadinya fotorespirasi ini. Singkatnya, mereka aka memodifikasi proses pengikatan CO2 agar tidak langsung berikatan dengan RuBP. Berdasarkan mekanisme modifikasinya, kita dapat membagi tumbuhan ke dalam dua kelompok, yakni tumbuhan C4 dan CAM. Sementara itu, tumbuhan yang langsung mengikat CO2 dengan RuBP disebut sebagai tumbuhan C3. Tumbuhan seperti padi, gandung dan kacang kedelai merupakan contoh tumbuhan C3. Siklus Calvin pada tumbuhan C3 terjadi secara umum pada sel-sel mesofil daun.
Proses Fotosintesis Pada Tumbuhan C4
Berbeda dari tumbuhan C3, pada tumbuhan C4 (jagung, tebu dan sorgum) terdapat sebuah proses pendahuluan yang terjadi sebelum Siklus Calvin, yakni proses yang disebut sebagai Hatch-Slack pathway yang terjadi di sel mesofil daun. Enzim PEP karboksilase mengikat CO2 yang diambil dari lingkungan dengan molekul beratom karbon tiga, yakni PEP dan menghasilkan molekul beratom karbon 4 (oxaloacetate). Molekul dengan empat atom karbon ini (pada Gambar 7 dicontohkan Malat) kemudian akan ditransfer ke sel bundle selubung (bundle-sheets cell) melalui plasmodesmata. Setelah sampai di bundle-sheets, malat kemudian akan melepaskan satu molekul CO2 dan satu molekul dengan tiga atom karbon (piruvat). Dengan menggunakan energi dari ATP, piruvat kemudian diubah menjadi PEP dan kembali melanjutkan siklus, sementara CO2 akan memulai Siklus Calvin. ATP yang digunakan dihasilkan oleh proses fotosintesisi siklik.
Bundle-sheets cell merupakan sel yang bersifat impermeable terhadap CO2. Artinya, CO2 yang telah masuk ke sini tidak akan dapat keluar sehingga apabila Hatch-Slack pathway terus berlangsung, konsentrasi CO2 di bundle-sheets terus meningkat. Perlu diketahui bahwa dalam memutuskan akan berikatan dengan O2 atau CO2, RuBP akan memilih mana yang konsentrasinya lebih tinggi. Mekanisme seperti ini tentunya sangat efektif untuk mencegah terjaidnya fotorespirasi. Bila mekanisme ini efekktif untuk mencegah fotorespirasi, mengapa tidak semua tumbuhan mengadaptasi Hatch-Slack pathway? Alasan yang pertama adalah jalur biokimia ini hanya ada pada tanaman yang nenek moyangnya mengalami mutasi yang menyebabkan adaptasi ini (yang ternyata menguntungkan bagi tumbuhan-tumbuhan C4). Tumbuhan tidak bisa begitu saja “memilih” jalur mana yang akan digunakan. Sistem fotosintesis yang mereka adaptasi sepenuhnya dipengaruhi oleh sejarah evolusi spesies mereka. Alasan kedua ialah Hatch-Slack pathway yang digunakan oleh tanaman C4 membutuhkan penggunaan ATP tambahan (yang diperlukan untuk konversi piruvat menjadi PEP). Dengan demikian, tanaman yang memanfaatkan jalur Hatch-Slack harus “membayar” energi tambahan ini, tidak seperti tumbuhan C3. Untuk itu, apakah jumlah energi tambahan yang harus dikeluarkan ini namun, di pabrik ini worth it juga menjadi pertimbangan. Untuk tumbuhan C4 sendiri, “biaya” tambahan ATP ini cukup worth it karena lokasi hidupnya yang kebanyakan di lingkungan kering.
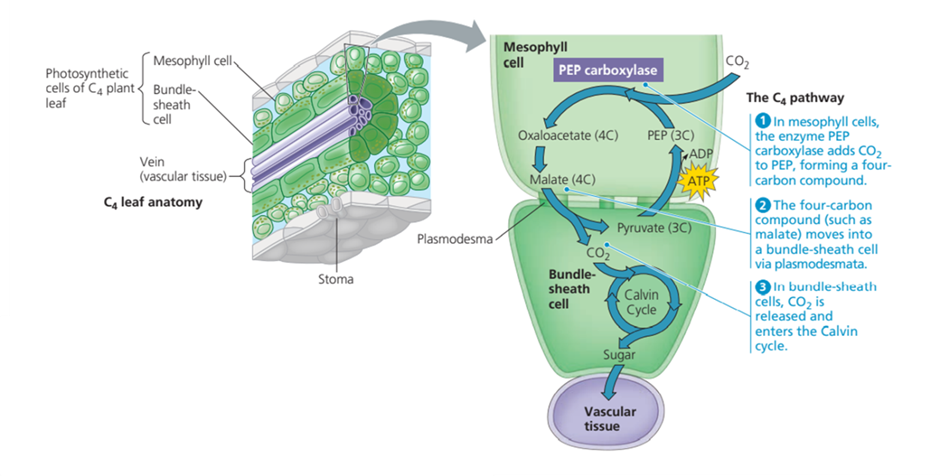
Gambar. Proses Fotosintesis Tumbuhan C4
Proses Fotosintesis Pada Tumbuhan CAM
Adaptasi fotosintesis CAM dapat dijumpai di banyak tanaman sukulen (penyimpan air), kaktus, nanas, dan beberapa famili tanaman lainnya. Tumbuhan ini membuka stomata pada malam hari dan menutupnya pada siang hari, kebalikan dari tumbuhan pada umumnya. Menutup stomata pada siang hari membantu tumbuhan menghemat air dan mencegah CO2 memasuki ke daun. Pada malam hari, ketika stomata terbuka, tanaman ini mengambil CO2 dan menggabungkannya ke dalam berbagai asam organik. Cara fiksasi karbon ini disebut sebagai crassulacean acid metabolism, atau CAM, diambil dari nama famili tanaman Crassulaceae, sukulen tempat proses tersebut pertama kali ditemukan. Sel-sel mesofil tumbuhan CAM menyimpan asam-asam organik yang mereka buat pada malam hari di dalam vakuolanya hingga pagi hari, ketika stomata menutup. Pada siang hari, ketika reaksi terang dapat memasok ATP dan NADPH untuk siklus Calvin, CO2 dilepaskan dari asam organik yang dibuat malam sebelumnya untuk dimasukkan ke dalam gula dalam kloroplas.
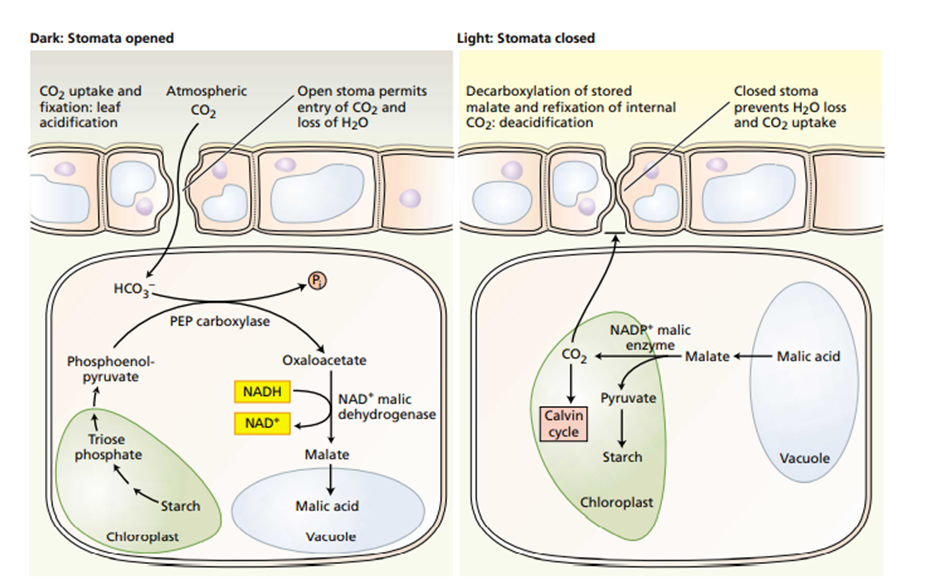
Gambar. Proses Fotosintesisi pada Tumbuhan CAM
Referensi
- Hopkins, W.G. & Hüner, N.P.A., 2009. Introduction to plant physiology 4th ed., Hoboken, NJ: John Wiley & Sons.
- Urry, L. A., Cain, M. L. 1., Wasserman, S. A., Minorsky, P. V., Reece, J. B., & Campbell, N. A. (2017). Essential biology. Eleventh edition. New York, NY: Pearson Education, Inc.
- Cooper, G. (2019) The Cell A Molecular Approach Eight Edition. Eight, Oxfore University Press. Eight. Oxford: Oxford University Press.
- Simon, E. (2015) Biology The Core. First Edit. Pearson Education.